KcsAチャネル:構造
この論文では、二つのドメインを含むStreptomyces lividansのカリウムチャネルであるKcsAに焦点を当てます。 最初は120残基を含む膜貫通部分である。 これについては、以下で詳細に説明する。 第二のドメインは、約40残基を含む細胞質ドメインである。 細胞質ドメインは、閉じた状態の安定性に寄与する。 これは,細胞質ドメインが結晶構造から除去されたときにチャネルの折り畳みと集合の効率の低下と熱安定性が影響することを示すデータに基づいて結論された。図5図2に見られるように、水で満たされたC末端細胞質ドメインは、細胞質に向かって直線的に延びる4ヘリックス束構造である。5-6Uysalによって生成された電子常磁性共鳴、EPR、データによって、C末端は、C末端ドメインが膜貫通ドメインを満たすところで膨らんだ伸張を有することが示されている。5この二つを接続する螺旋の伸張は、チャネルの開口部の間に役割を果たす可能性のある他の部分よりも高い程度の柔軟性を有する。5C末端ドメインの機能を完全に理解するためには、まだ多くの研究が必要です。 ドメインの提案された機能は、それが正体不明のプラスミックアクチベータータンパク質またはリガンドの受容体として役立つかもしれないということであるが、他の人はイオンの透過経路に有意な影響がないと主張している。 構造はまだ任意の側鎖の配向を含んでいないが、ヘリックスの配置は、サブユニット内の静電相互作用を示唆しています。図6図3は、細胞質ドメインが開いたチャネルと閉じたチャネルとの間で二つの細菌チャネルを比較する方法を示しています。7論文の残りの部分は、カリウムを選択する膜貫通ドメインに焦点を当てます。
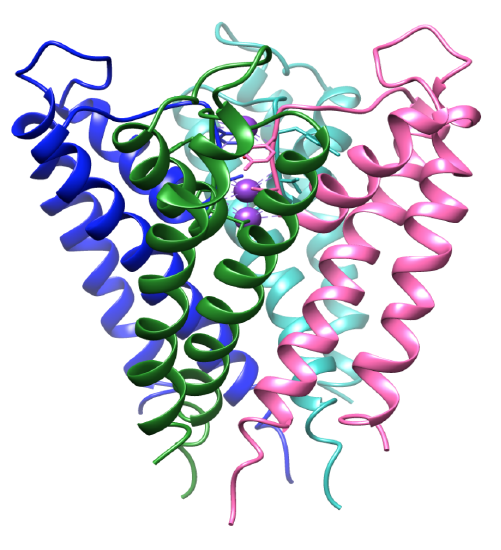
図3. 膜貫通ドメイン。 それぞれのサブユニットは異なる色をしており、カリウムイオンは紫色をしています。 PDB1BL8.
カリウムイオンの選択性は、カリウムチャネル選択性フィルターの構造に大きく依存しています。 チャネルの配位環境は、カリウムイオンに特異的に結合する選択的配位子を有する。8カリウムチャネルタンパク質は四量体であり、中央孔を形成する四つの同一のサブユニットを含む。8-9イオンは開閉する気孔に流れ、イオンを中央キャビティに指示します。 このサブユニットには2つのαヘリックスが含まれており、1つは中央の細孔、内側のヘリックス、もう1つは脂質膜に面しており、外側のヘリックスと呼ばれています。内側および外側の螺旋と共に、各サブユニットは、細孔螺旋を含む。3,8図3に示すように、サブユニットは傾いており、選択性フィルターは膜の細胞外表面の近くにある。 四つの内側ヘリックスは、それらが細胞内表面の近くに一緒に来る方法で構造化されています。 内部および細胞外の通路は酸性アミノ酸によって否定的に荷電されます。 これは膜の近くで陽イオンのより高い濃度および反対の充満のためにより少ない陰イオンを可能にします。8

図4. カリウムチャネル(Streptomyces lividansからのKcsA構造)には、主要な構造成分が標識されています。 注意すべき主な側面は、細孔、空洞、および選択性フィルタ(SF)である。 これらは、カリウムイオンが通過するチャネルの部分である。 第一に、イオンは細孔を通って流れ、細孔が開いているときに水和された空洞に入る。 その後、カリウムイオンは脱水され、SFを通過して細胞に入るように選択される。 この画像には、視覚化を容易にするために2つのサブユニットのみが表示されていることに注意してください。 PBD1K4C.
細孔領域は、細孔を遮断するサソリ毒素で最初に同定された。 それらは妨害および機能障害を引き起こす気孔の入口でアミノ酸と相互に作用しています。 細孔は、図4に示すように、カリウムイオンが入る場所であり、そこでそれが中央空洞に移される。 この時間の間にカリウムイオンは水和させてとどまります。8細胞内孔は、カリウムイオンが開いているときに細胞に入ることを可能にするチャネルの特徴であり、または閉じたときに入ることを遮断する。 チャネルは膜の潜在性に変更があるとき開き、キャビティフィルターにカリウムイオンの流入を許可する。 従って選択的な結合が選択性フィルターで起これば、潜在性は膜を渡る低下しま、気孔を閉めます。1気孔およびキャビティはhydrophobically並びま、静電気の障壁を下げる機能を許可します。 より低い静電気の障壁がある必要があります従って脱水の精力的な費用はイオン浸透(誘電性の障壁)のための大きいエネルギー障壁が償われた原因で 従って、水によって満たされるキャビティは精力的に好ましい環境を可能にする。3,8,10,11
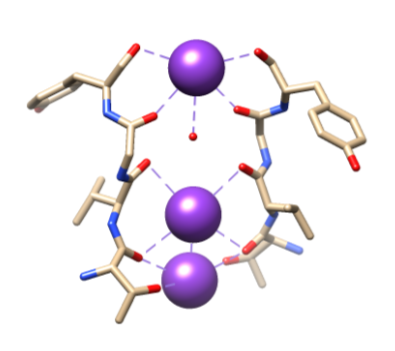
図5. Thr75、Val76、Gly77、およびTyr78の骨格Oによる特定の配位子へのカリウムイオン(紫色)の結合。 視覚化を容易にするために、ここでは完全な調整は示されていません。 Samsomから再生されました。11
次に、アミノ酸に属する極性主鎖原子が並ぶ選択性フィルター(SF)にイオンを選択する必要があります。 SFは非常に狭いので、イオンが入る前にまず脱水しなければなりません。 SFには2つの本質的な特徴があります。 まず、主鎖原子は、脱水カリウムイオンをスタッキングすることによって配位するのに適した寸法を作り、連続した酸素リングを作る。 これはフィルターを通って移動している間カリウムイオンが各場所に達するために移動しなければならない非常に小さい間隔を作成する。 これにより、図6に示すように、四つの立方八重配位サイトのシーケンスが形成されます。3,8いつでも、これらのサイトは2つのK+イオンと2つの水分子が交互に配列して占有されています2つ目の特徴は、図5.9に示すT-V-G-Y-Gのシーケンスに続くSFの周りのタンパク質パッキングです。2つのカリウムイオンがSFに入ると伝導が起こり、2つのカリウムイオンの間で起こる反発力とSFからイオンへの引力のバランスを引き起こします。8この反発はカリウムイオンが結合場所に堅く結合のための本質的な類縁を克服するのを助けます。3選択メカニズムのさらなる議論は、以下に議論される。
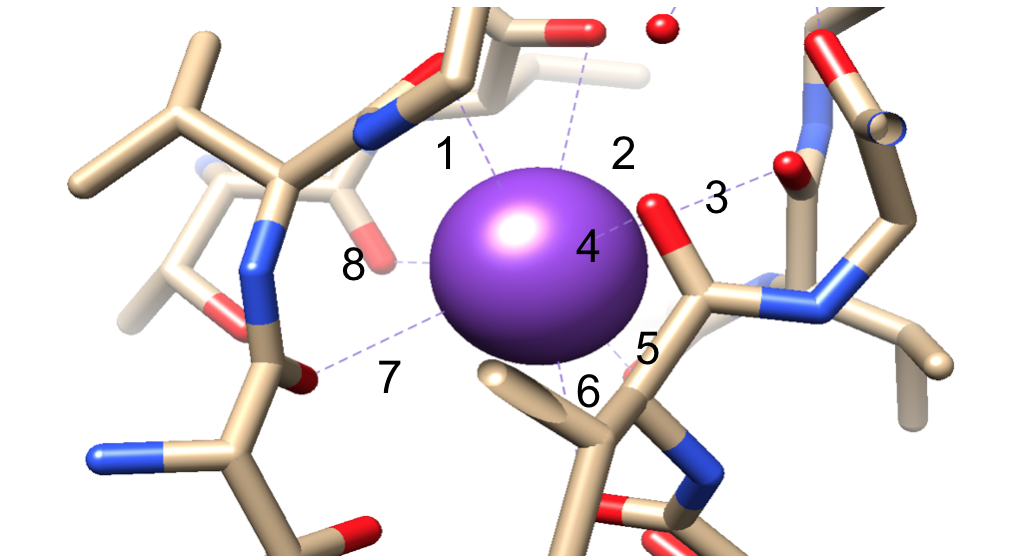
図6. 八重配位部位におけるカリウムイオン(紫色)の一例。 より容易な視覚化のために番号を付けられる結合。