DISKUSE
zjistili Jsme, tady genomu o velikosti 25 druhy pakomárů; tyto hodnoty byly shodné s velikostí genomu ze tří druhů dříve reportedin literatuře (Petitpierre, 1996; Schmidt-Ott et al., 2009), které byly složené mezi 0,13 a 0,21 pg DNA. Výsledky této studie ukázaly, že velikosti genomu chironomidů byly rovnoměrně velmi malé (obr. 3, Tabulka 1). Získaná střední hodnota C pro čeledi Chironomidae je 0,13 + 0.03 PG DNA, která je zřetelně menší než velikost genomu d.melanogaster. Nejmenší velikost genomu u hmyzu byl zaznamenán v Strepsiptera Caenocholax fenyesi s o 0.11 pg DNA (odpovídající zhruba na 108 Mb), i když Hessian létat Mayetiola destruktor bylo také navrhl, že mají ještě menší velikost genomu 0,09 pg (Johnston et al., 2004; Gregory, 2014). Zde jsme se hlásí nejméně tři druhy s menší genomy než jakýkoli známý hmyz: C. tsushimensis, Diamesa japonica a Hydrobaenus tsukubalatus, s C-hodnoty 0,07 pg, 0,08 pg a 0,08 pg, v tomto pořadí (Tabulka 1). Pokud je nám známo, C. tsushimensis (obr. 1B) lze nyní považovat za nejmenší velikost genomu jakéhokoli známého hmyzu, s hodnotou C 0,07 PG DNA, což odpovídá zhruba 68 Mb. Tato hodnota je shodná s velikosti genomu z příbuzných druhů, Clunio marinus, který byl odhadován jednou až 95 Mb a více nedávno 87.2 Mb (Kaiser a Heckel, 2012; Tobias S. Kaiser, osobní sdělení).
nedávno byl sekvenován genom antarktického midge, B. Antarktida (Kelley et al., 2014). Autoři tvrdili, že velikost genomu tohoto druhu se odhaduje na 89.5-105 Mb, byl nejmenší ze všech hmyzu. Zde však ukazujeme, že tato velikost genomu je obsažena v normálním rozmezí hodnot chironomidů. Autoři také navrhli, že malá velikost genomu B. Antarktida byla adaptací na extrémně chladné prostředí. Naše data mohou tuto hypotézu potvrdit, protože d. japonica, alpský druh z podčeledi Diamesinae, také představuje malou velikost genomu. Další příbuzný Diamesinae, Himálajský midge Diamesa sp. bylo zjištěno, že je aktivní při teplotách až -16°C (Kohshima, 1984). Nicméně, Telmatogeton japonicus, který je také schopen vyvinout při velmi nízkých teplotách (Danks, 1971; Sunose a Fujisawa, 1982), ukazuje, poměrně velké velikosti genomu, ve srovnání s pakomárů rodinný průměr. Zde je třeba vzít v úvahu fylogenetický vliv na velikost genomu, protože b. Antarktida patří do podčeledi Orthocladiinae, která v průměru představuje malé velikosti genomu. Druhy C. tsushimensis a Hydrobaenus tsukubalatus, zobrazující nejmenší velikost genomu mezi chironomids, také patří do podčeledi Orthocladiinae. Tedy malá velikost genomu B. Antarktida může jednoduše představovat rodový rys, nikoli výsledek extrémní adaptace na chlad.
P. vanderplanki je jediný druh hmyzu známo, že dosáhnout anhydrobiosis ve zcela dehydrované stavu (Obr. 1D) a zotavit se po rehydrataci (Hinton, 1951, 1960a; Watanabe, 2006; Cornette a Kikawada, 2011). Naše výzkumná skupina nedávno zveřejnila srovnávací analýza genomu sekvence sušení-tolerantní Polypedilum vanderplanki a vysoušení citlivé podobných skupin Polypedilum nubifer (Gusev et al., 2014). Srovnání ukázalo, že oba druhy měly podobné velikosti genomu a že pouze omezená sada duplikovaných genových shluků souvisela s anhydrobiózou U P. vanderplanki (Gusev et al ., 2014). Genom velikostí odvodit z genomu shromáždění byly 104 Mb pro P. vanderplanki a 107 Mb pro P. nubifer a tyto hodnoty se výrazně liší od genomu velikostí naměřených u těchto druhů v této studii, což odpovídalo přibližně 98 Mb. P. vanderplanki a P. nubiferovy genomy vykazovaly podobně nízký podíl opakování DNA a bylo nalezeno pouze malé množství transpozovatelných prvků ve srovnání s jinými druhy dipteranů. Toto snížení podílu transponovatelných prvků bylo také pozorováno v antarktické midge, B. Antarktida (Kelley et al ., 2014). Průměrná intron délka byla také výrazně snížena v Antarktidě midge a v obou P vanderplanki a P. nubifer stejně (Gusev et al., 2014). Vzhledem k tomu, vysychání citlivé a zmrazení netolerantní P. nubifer akcie těchto funkcí s Antarktickou midge a anhydrobiotic midge, nízký podíl transpozonů a krátké intron délka může být charakteristika, kterou sdílí všechny chironomids a může být přímo souvisí s tolerancí k extrémnímu prostředí, i když adaptivní účinek nemůže být vyloučen.
obr. 3.
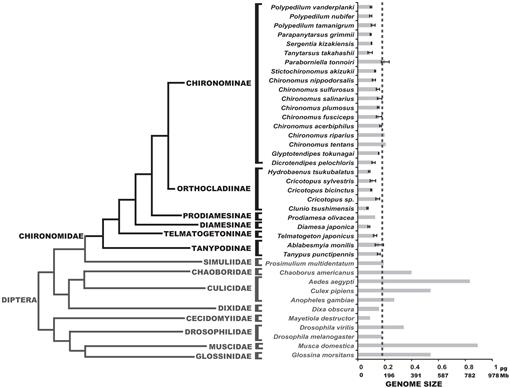
fylogeneze Diptera vykazující odhadované C-hodnoty. Rodina chironomidů je zobrazena černě. Pomlčka ukazuje velikost genomu d. melanogaster. Fylogenetický kladogramu je konsensus z různých Pakomárů a Dipteran stromy (Yeates a Wiegmann, 1999, 2005; Saether, 2000; Cranston et al., 2012). Sloupce chyb ukazují směrodatnou odchylku. Velikosti genomu bez chybových pruhů byly získány z předchozích studií uvedených v databázi velikosti genomu zvířat (Gregory, 2014).
obr. 4.
Vztah mezi velikostí těla a velikosti genomu mezi druhy pakomárů zkoumány v této studii. A) pro každý druh je průměrná celková délka těla uvedena v milimetrech a průměrná hodnota C je vyjádřena v PG DNA. Mezi oběma proměnnými byla pozorována pouze slabá pozitivní korelace (r2 = 0, 171, n = 25), materializovaná pevnou čarou. (B) na úrovni rodu byly prostředky stejných hodnot transformovány metodou fylogeneticky nezávislých kontrastů založených na fylogenezi Cranston et al. (2012). Korelace mezi těmito transformovanými kontrasty, materializovanými plnou čarou, nebyla významná (r2 = 0,118, n = 13). Analýza dat byla provedena na Prism 4.0 software pro Macintosh (GraphPad Software, Inc.).

Co by mohlo být adaptivní rysy potenciálně spojené s tiny velikost genomu u Pakomára rodinu? Ve skutečnosti bylo navrženo několik vývojových a ekologických faktorů, které korelují s velikostí genomu. Za prvé, pozitivní korelace mezi velikostí genomu a velikostí těla byla nalezena v mnoha taxonech a tato korelace se jeví jako relativně obecný jev (Gregory, 2005). Tato korelace byla hlášena také u komárů (Ferrari a Rai, 1989), ale to není vždy jasné, v jiné hmyzí taxony (Gregory a Hebert, 2003; Ardila-Garcia a Gregory, 2009). V rámci chironomidů jsme našli pouze slabou pozitivní korelaci mezi velikostí genomu a velikostí těla (obr. 4A). Průměrná hodnota C za rodu Chironomus (0.15 pg) byla vyšší než u rodu Polypedilum (0.10 pg), a tento rozdíl může souviset s obecně větší velikost těla Chironomus druhů. Fylogenetická vzdálenost mezi oběma rody by však také mohla vysvětlit tento rozdíl velikostí genomu. Následně jsme opravili naše data na úrovni rodu fylogeneticky nezávislou analýzou kontrastů (Garland a Adolph, 1994; Garland et al., 2005) a v důsledku toho se korelace mezi velikostí genomu a velikostí těla ukázala jako nevýznamná (obr. 4B). Úzké rozpětí Pakomárů, že velikost genomu (pouze dva—trojnásobně) a průměrné malé velikosti těla těchto druhů (většinou nepřesahuje několik milimetrů) pravděpodobně těžké najít nějaké korelace mezi oběma znaky v Pakomára rodiny.
u hmyzu byla navržena další korelace mezi velikostí genomu a vývojovou složitostí. Vzhledem k tomu, že hemimetabolous hmyzu, s postupným nymfí líná jen, ukázat širokou škálu velikostí genomu (C-hodnoty od 0.18 do 16.93 pg), holometabolózní hmyz s jejich komplexní metamorfózou má hodnoty C omezené v domnělém Prahu 2 pg, pro většinu z nich (Gregory, 2002, 2005). Vlivem vývojové složitosti na velikosti genomu je zde zřejmé, protože holometabolous insects podstoupit intenzivní morfologické přestavby během omezené doby proměny a malá velikost genomu představuje výhody pro dosažení mobilní divize vysokým tempem. Chironomidy jsou holometabolózní hmyz a jejich metamorfóza je extrémně rychlá. Jejich pupální fáze může trvat pouze jeden den, nebo dokonce jen několik hodin u některých druhů (Cranston, 1995c). Například C. tsushimensis vykazuje komplexní metamorfózu s výrazným sexuálním dimorfismem (obr. 1B).
vztah mezi velikostí genomu a rychlostí buněčného dělení také ovlivňuje rychlost vývoje. Například rychlý životní cyklus mšic byl navržen tak, aby souvisel s jejich malými velikostmi genomu (Ma et al., 1992). Malé velikosti genomu by tedy měly představovat výhodu pro druhy chironomidů, které se rychle vyvíjejí v dočasném prostředí. Chironomids z rodu Clunio může vyvinout dočasné příliv bazény a také ukazují synchronizované vznik dospělých ve vztahu k měsíční cyklus a během krátké okno odlivu (Kaiser a Heckel, 2012). K dosažení tohoto cíle by měla být zapotřebí vysoká míra vývoje. Všimněte si, že malé mnohoštětinatec kroužkovců rychle rozvíjí v podobné intersticiální prostředí ukazují menší velikost genomu než macrobenthic druhů (Gambi et al., 1997). Pro antarktické midges, jako je B. Antarktida nebo Eretmoptera murphyi (Lee et al., 2006; Worland, 2010), vývoj larev trvá dva roky, většinou zastaven ve zmrazeném stavu, ale metamorfóza a reprodukce musí nastat během velmi krátkého letního období. V tomto případě je zapotřebí vysoká míra vývoje, což může usnadnit malý genom B. Antarktida. Tento jev dokládá, že studie o krytosemenných rostlin, ukazuje, že tento druh schopen dokončit celý jejich životní cyklus během krátkého Antarktického léta představila nejmenší velikost genomu (Bennett et al., 1982). Další extrémní, anhydrobiotic midge P. vanderplanki je přizpůsoben pomíjivé rock pools. I zde může být vysoká míra vývoje v dočasném prostředí spojena s malou velikostí genomu tohoto druhu. Australský druh P. tonnoiri, který je také vystaven vysychání na podobných skalních bazénech (Jones, 1975; Adams, 1985; Frouz et al., 2003), ukazuje relativně velkou velikost genomu pro chironomid (obr. 3 a tabulka 1).
abiotické stresy také pravděpodobně ovlivňují integritu DNA a v důsledku toho velikost genomu. Například larvy P. vanderpianki zažívá masivní poškození DNA po anhydrobióze a účinná Oprava DNA nastane během několika dnů po rehydrataci (Gusev et al., 2010). U rostlin byla zjištěna negativní korelace mezi velikostí genomu a tolerancí záření (Bennett a Leitch, 2005) a míra mutace byla také nižší u druhů s menší velikostí genomu (Abrahamson et al ., 1973). Z toho vyplývá, malá velikost genomu P. vanderpianki může představovat výhodu, aby se zabránilo hromadění mutací během opravy DNA události spojené s cykly vysychání a rehydrataci. Jako mořský druh je C.tsushimensis také vystaven stresu s vysokou slaností, který je smrtelný pro většinu druhů hmyzu. Je známo, že vysoká slanost a další abiotické stresy vytvářejí intracelulární reaktivní kyslíkové druhy (ROS), které byly navrženy jako hlavní zdroj poškození DNA (Franca et al ., 2007; Gill and Tuteja, 2010). To může také souviset s malou velikostí genomu. Jiné mořské druhy tolerantní k vysoké slanosti, jako je T. japonicus nebo Chironomus salinarius, však nevykazují velikosti genomu tak malé jako C. tsushimensis. Druhy odolné vůči kyselinám, jako je Chironomus sulfurosus, Chironomus acerbiphilus nebo Polypedilum tamanigrum (obr. 1C) může přežít v horkých pramenech s pH tak nízkým jako 1,4 (Doi et al ., 2004; Takagi a kol., 2005). Zde opět, žádná jasná korelace mezi tolerance na kyselé stres a malé velikosti genomu by mohl být pozorován a fylogenetický vliv se zdá důležitější, s menší velikost genomu u rodu Polypedilum a větší velikost genomu v rodu Chironomus.
Na závěr, malá velikost genomu pozorovány v chironomids jsou pravděpodobně výsledkem rodové snížení počtu transpozonů a délka intronů v jejich genomu struktura. U čeledi Chironomidae se velikost genomu lišila podle fylogeneze na úrovni podčeledi a na úrovni rodu. Jak odvodit z korelace v různých taxonů, malá velikost genomu z chironomids teoreticky mohl představuje předběžná adaptace na nestabilní a extrémní prostředí prostřednictvím vysoce vývojové rychlost a nízkou mutační rychlostí. Doufáme, že tato studie bude podporovat budoucí Pakomárů genomu projekty a komparativní genomika v rámci tohoto taxonu pomůže pochopit, jak Pakomárů genomy jsou v interakci s jejich prostředím.