Mikrobiyhteisörakenteeseen vaikuttavien tekijöiden analyysi
ympäristötekijöiden ja yhteisön varianssin välisten mahdollisten suhteiden selvittämiseksi luotiin RDA-analyysi (Kuva. 7). Yhteensä kahdeksan ympäristötekijää, mukaan lukien hivenaineet Fe, Co, ni, lämpötila, suolaisuus, syvyys, kosteus ja RO. Näytteenottopaikalta mitattiin syvyys ja säiliön lämpötila, ja muita tietoja saatiin paikallisten kaivosten geologisista tiedoista (KS.Taulukko 2). Tiedot esitetty kuvassa. 7a paljasti, että tässä tutkimuksessa löydettyihin bakteeriyhteisön koostumuksiin vaikuttivat merkittävästi Fe, Ni, Kosteus, suolaisuus ja RO. Kaikki yhteisöt, lukuun ottamatta C4, C7, C10, korreloivat positiivisesti RO: n kanssa; C4, C7 ja C10 positiivisesti korreloivat Fe: n, Ni: n ja kosteuden kanssa. Koentsyymi m-metyylitransferaasi vaatii Co: ta, joka on tärkeä entsyymi metanogeenien biokemiallisessa aineenvaihdunnassa ; siksi Co: n vaikutus arkaaliyhteisöön on suurempi kuin bakteeriyhteisön. Alkuaineet Fe, Co, ja ni; sekä kosteus, näyttivät olevan merkittävimmät ympäristötekijät, joita seurasi ro ja suolapitoisuus arkaaisessa yhteisössä. Co-ja C1 -, C8 -, C9-ja C10-yhteisöjen välillä on merkittävä positiivinen korrelaatio. Kaikki yhteisöt paitsi C2, C4, C6 ja C5 korreloivat negatiivisesti suolapitoisuuden kanssa (kuva. 7b).

RDA (redundancy analysis), joka perustuu bakteerien (A) ja archaea (b) tasoon hiilikerroksen ympäristötekijöillä ja hiilen ominaisuuksilla. Iskukertoimen pituus on pidempi, vaikutuksen vaikutus on suurempi, ja vastaavasti, kun iskukerroin on lyhyempi, iskun vaikutus on kevyempi. Kun ympäristötekijä on akuutisti kulmassa näytteeseen, on positiivinen korrelaatio, ja kun ympäristötekijä ja näytteen kulma ovat tylsiä, on negatiivinen korrelaatio
kivihiilen sijoitus
kivihiilen ”hyppy” viittaa geologisen historian aikana tapahtuneisiin fysikaalisiin ja kemiallisiin muutoksiin hiilen lämpötilassa ja paineessa. Kivihiili on siis käynyt läpi asteittaisen ja äkillisen muutoksen prosessin. Neljä hyppyä vastaa RO: ta 0,6, 1.3, 2, 5 ja 3, 0%. Riippumatta tarkasteltavasta arkaalisesta tai bakteeriyhteisöstä kivihiilen sijoitus vaikuttaa jonkin verran bakteerien monimuotoisuuteen ja runsauteen. Kun kivihiilen sijoitus kasvaa sekä arkaaisissa että bakteerikunnissa, yhteisön monimuotoisuus osoittaa tiettyä laskevaa suuntausta yleisesti (Kuva. 8). Lisäksi mikro-organismit voivat vaikuttaa hiilen koostumukseen, jota hiiliryhmät säätelevät. Keski-ja matalarankaiset hiilet sisältävät suuria määriä kasvien kehittyneitä aineita ryhmässä 1, joka sisältää paljon kasvien kehittyneitä aineita. Siellä on enemmän vetyä, happea ja typpeä, ja bakteerien tarvitsemat ravinteet ovat runsaita. Kivihiilituotannossa orgaaniset aineet tuottavat paljon kosteutta ja nestemäisiä hiilivetyjä. Samalla myös hiilen sisältämän vedyn ja hapen sivuketjut ovat runsaat. Nämä nestemäiset ja kiinteät aineet muodostavat bakteereille elämän perustan. Tämän vuoksi vetyä tuottavien bakteerien ja metanogeenien runsaus ja monimuotoisuus kivihiilessä on tällä alueella suhteellisen suuri. Ro: n lisääntymisen myötä hiilen vedyn ja hapen sivuketjupitoisuus vähenee rajusti ja pieneliöiden käytettävissä olevat komponentit vähenevät. Siksi bakteeri-ja arkaaliyhteisöjen lajirunsaus ja monimuotoisuus ryhmässä 2 ja ryhmässä 3 vähenee kaiken kaikkiaan. Tähän mennessä luonnossa on havaittu biogeenisen hiilivetyetaanin heijastuskyky olevan 2,0% (C4 Hebi). Ro > 2,5%: n jälkeen pieninä molekyyleinä muunnettavissa olevat orgaaniset yhdisteet ovat olleet hyvin harvinaisia, mutta ryhmässä 4 on ollut enemmän monimuotoisuutta ja runsautta. Spekuloimme, että pohjaveden tuomat ravinteet ovat tällä hetkellä käytettävissä bakteerien lisääntymiseen. Pohjaveden eri alueilla ja eri vuodenaikoina tuomat ravinteet ovat saattaneet edistää lajien monimuotoisuutta. Yksi syy ryhmän 4 kasviston suurempaan monimuotoisuuteen voi olla se, että C8 Jiaozuo Jiulishanin alueella on paremmat pohjaveden valumaolosuhteet ja vahvempi latautuminen. Se voi kuljettaa ravinteet kasvistoon, joten monimuotoisuus ja runsaus ovat suurempia kuin ryhmä 2. On syytä huomata, että arkaaisten yhteisöjen monimuotoisuus ja runsaus korreloivat jossain määrin negatiivisesti kivihiilen kanssa. Bakteerikuntien lajirunsaus korreloi kuitenkin positiivisesti kivihiilikantojen kanssa, ja monimuotoisuus osoittaa laskevaa suuntausta. Kivihiililuokkien nousun myötä jotkut bakteeriryhmät sopeutuivat vähitellen eri kivihiililuokkien ympäristöön ja voivat kasvaa ja lisääntyä suurina määrinä, ja metanogeeneja on vaikea sopeutua kivihiililuokkiin.
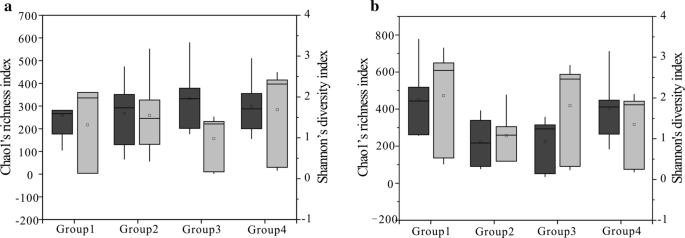
Chao1: n (Tummanharmaa) ja Shannonin (vaaleanharmaa) indeksi neljälle ryhmälle (Hiilinäytteet jaettiin neljään ryhmään Ro: n arvon mukaan bakteeriyhteisölle (a) ja archaea-yhteisölle (b), ryhmä 1 edustaa arvoa alle 0,6%, ryhmä 2 on arvo välillä 0,8-1,1%, ryhmä 3 on arvo välillä 1,4-1,8%, ryhmä 4 edustaa arvoa välillä 2,67-3,15%), joka on johdettu alueista. 25. ja 75. prosenttipisteet ilmoitetaan laatikoiden ulkoreunoilla, kun taas enimmäis – ja vähimmäisarvot esitetään viiksien päillä ja mediaani vaakasuoralla viivalla kussakin laatikossa.
hivenaineet
hivenaineet voivat edistää mikro-organismien kasvua tietyllä alueella, jossa solu ylläpitää alkuaineiden homeostaasia aineenvaihdunnan säätelyn avulla. Hivenaineita voi esiintyä myös erilaisissa entsyymeissä, joita mikro-organismit voivat absorboida ja käyttää anaerobisen aineenvaihdunnan prosessissa, jolla on vaikutusta vetyä tuottavien bakteerien ja metanogeenien yhteisörakenteeseen (Taulukko 3).
Fe: llä ja Ni: llä on suurempi vaikutus vetyä tuottaviin bakteereihin kuin Co: lla . Fe ja Ni voivat osallistua hydrogenaasien ja muiden metalloentsyymien synteesiin ja aineenvaihduntaan mikro-organismeissa. Kun Fe: n ja Ni: n pitoisuus kasvaa tietyllä alueella, kasvaa myös vetyä tuottavien bakteerikantojen runsaus ja monimuotoisuus. Fe: n ja Ni: n pitoisuus C7: ssä on paljon korkeampi kuin muilla alueilla, ja tässä työssä on havaittu, että Clostridium on vetyä tuottava bakteeri. Tämä havainto osoittaa, että hivenaineiden liiallisilla pitoisuuksilla voi olla myrkyllinen vaikutus mikro-organismien kasvuun ja ne estävät metalloentsyymien toimintaa. Fe: n tasot C4: ssä, C6: ssa ja C9: ssä eivät poikenneet merkittävästi toisistaan ja olivat stabiileja 3500 mg Kg−1: ssä (kuva. 9). Ni: n suhteellinen runsaus näillä kolmella alueella on C6 > C9 > C4, mikä vastaa runsausjärjestystä (myös C6 > C9 > C4), mutta monimuotoisuusjärjestys on C6 > C4 > C9. Clostridium -, Klebsiella -, Enterobacter-ja Citrobacter-sukujen jäseniä havaittiin C4 -, C6-ja C9-yhteisöissä, mukaan lukien ne, joiden pitoisuudet ja monimuotoisuustasot olivat suuremmat kuin muilla alueilla.

Fe: n, Co: n ja Ni: n hivenainepitoisuus kivihiilinäytteissä
arkaaisessa yhteisössä Fe: n, Co: n ja Ni: n vaikutus metanogeeneihin on vielä tärkeämpi. Co on keskeinen tekijä metanogeenisen koentsyymin F430 synteesissä, ja kärkikolmikon Co-pitoisuus on C8 > C7 > C10, metanogeenilajit ja runsaus on C8 > C7 > C10. Co-pitoisuus korreloi positiivisesti metanogeenien runsauteen ja monimuotoisuuteen jossain määrin. Vaikka Fe: n pitoisuus C7: ssä on paljon korkeampi kuin muilla alueilla, se ei vaikuta metanogeenien jakautumiseen alueella. Metanogeeneja, jotka voivat sisältää Fe: tä, on vain muutamia tyyppejä—aiemmissa tutkimuksissa löydettiin vain yksi laji, nimeltään Metanothermobacter. Monoferrinen hydraasi M. marburgensis-organismin metanogeeneissa katalysoi metenyyli-H4MPT+: n ja H2: n reversiibeliä reaktiota metyleeni-h4mpt: n ja H+: n muodostamiseksi; metaanin tuottamiseksi CO2: sta ja H2: sta . Myös hydrotrofista metaboliaa käyttävät metanogeenit voivat sisältää samankaltaisia entsyymejä. Lisäksi suurella osalla metyylitrofisista metanogeeneista on myös arveltu olevan tällaisia entsyymejä Metanoculleus-ja Metanobakteereita lukuun ottamatta. On arveltu, että Metyylitrofisissa metanogeeneissa voi olla FE: hen liittyvä metalloentsyymi.
Pohjavesiolosuhteet
pohjavesi luo suoraan tai välillisesti ekologisen perustan ekstremofiilien kasvulle ja aineenvaihdunnalle kivihiilisaumassa. Toisaalta pohjavesi lataa runsaasti ravinteita bakteeri-ja arkaaliyhteisöille; toisaalta pohjaveden ympäristöolosuhteet (Eh, pH, suolaisuus, ionikoostumus ja hivenaineet) vaikuttavat suoraan mikrobien kasvuun ja metabolisen entsyymin toimintaan. Pohjaveden ympäristöolosuhteet liittyvät suoraan kivihiilen käyttöön ja hajoamiseen, ja kivihiilisaumassa sijaitsevilla mikro-organismeilla on erilaiset yhdyskuntarakenteet ja toiminnalliset ominaisuudet.
Mikrobiravinnesubstraatit ovat yleensä liuenneita. Kaivosalueen valumavyöhyke voi mahdollistaa hiilisaumojen säilymisen. Erittäin läpäisevillä säiliöillä on positiivinen vaikutus vetyä tuottavien bakteerien ja metanogeenien kasvuun ja lisääntymiseen, kun taas metamorfismilla on merkittävä negatiivinen vaikutus hiilen läpäisevyyteen hiilivarastoissa . Alueilla, joilla on biogeeninen CBM, C2 -, C4 -, C6-ja C7-yhteisöt on kaikki dokumentoitu hyvin. Nämä yhteisöt kuuluvat matala-ja keskihiilen luokkaan, hiilen huokoisuus on suhteellisesti korkeampi kuin korkea-arvoisen hiilen, pohjavesi voi antaa ravinteita kivihiilisauman mikrobeille ajoissa. Nykyinen CBM kehitysvyöhyke Powder – joen valuma-alueella Yhdysvalloissa. on keskittynyt pääasiassa pohjaveden valumavyöhykkeelle. C6-kaivosalueella sijaitsevasta matalasta CBM-kaivosta saadut kaasun stabiilit isotooppitiedot vahvistivat myös biogeneettisen CBM: n esiintymisen alueella. Toisen syvän CBM: n kaasun stabiilit isotooppitiedot osoittivat kuitenkin hyvin, että CBM on pääasiassa termogeeninen. Tulokset osoittavat, että hautauksen syvyyden kasvaessa valumaolosuhteet heikkenevät ja mikro-organismin ravinteiden kuljettaminen vaikeutuu, mikä johtaa yhteisön runsauden ja monimuotoisuuden vähenemiseen. NO: n katto ja lattia. 2 hiilisaumassa C2-alueella on suhteellisen vakaita mutakivi-ja savikivikerroksia, mikä vaikeuttaa hiilisaumassa olevien vetyä tuottavien bakteerien ja metanogeenien nestemäisten ravinteiden saantia ja rajoittaa niiden kasvua ja aineenvaihduntaa ja siten niiden yhteisöllistä monimuotoisuutta ja runsautta. Huomaa, että tällä alueella chao1-indeksi on 240 ja Shannonin indeksi on 1,38 bakteerikunnassa. Metanogeenien chao1-indeksi on 82, Shannonin indeksi on 0,56. Hiekkakivi halkesi pohjavesialueella katto C4 alueella No. 21 hiilen sauma on parempi ladata olosuhteet ja täyttää hiilen sauma vedellä. On mahdollista, että mikrobiyhteisö kokee kumulatiivisia vaikutuksia eri ravintoaineiden riittävästä saatavuudesta, joka vaikuttaa kuljetukseen verrattuna C2-alueen mikrobiyhteisön runsauteen ja monimuotoisuuteen, joka on parantunut huomattavasti. Tällä alueella vetyä tuottavien bakteerien Chao1-indeksi on 148, Shannonin indeksi on 1,52, metanogeenien Chao1-indeksi on 368 ja Shannonin indeksi on 2,35. C6-alueen hiekkakivestä halkeava akviferi on suora veden täyttämä akviferi, jonka hiilisauma on nro 3. Murtumien kehittyminen kerroksen sisällä ja kohtalainen akveositeetti vaikuttavat myös aktiivisesti yhteisön runsauteen ja monimuotoisuuteen. Tässä vetyä tuottavien bakteerien Chao1-indeksi on 472, Shannonin indeksi on 1,56, metanogeenien Chao1-indeksi on 384 ja Shannonin indeksi on 1,08. Näin on myös C7-alueella, No. 5-hiilisaumalla on suoraan murtunut akviferi, jolla on hyvät latautumisolosuhteet, vetyä tuottavien bakteerien runsausindeksi on 458, Shannonin indeksi on 1.98; metanogeenien chao1-indeksi on 256 ja Shannonin indeksi 2,47. Siksi vetyä tuottavien bakteerien ja metanogeenien lajidiversiteetti C4: ssä, C6: ssa ja C7: ssä on suurempi kuin C2: ssa.
pohjaveden ympäristöolosuhteet vaikuttavat suoraan mikro-organismien kasvuun ja aineenvaihduntaan. Kivihiilipohjaisen pohjaveden pH-arvo on yleensä neutraali, mutta pH−arvo vaihtelee välillä 6,5-8,4 hiekkakivestä murtuneessa pohjavesialueella C4-alueella nro 21 kivihiilisauma ja suolapitoisuus on 1,0 g L-1. Vuonna suora pohjavesialueella kerros No. 3 hiilisauma C6-alueella, pH vaihtelee välillä pH 6,8-8,0 ja suolapitoisuus on 0,7 g L−1. C7-alueen pohjaveden pH-arvo on 6,1−7,3 ja suolapitoisuus 1,25 g L-1. PH-arvo C4: ssä, C6: ssa ja C7: ssä on lähellä neutraalia ja mineralisaatioaste on alhainen, jossa mikro-organismiyhteisöllä on parempi kasvu, suurempi runsaus ja suurempi monimuotoisuus. Lisäksi pohjaveden suolaisuus ja ionikoostumus liittyvät läheisesti hiilisauman anaerobiseen pelkistysympäristöön. Esimerkiksi SO42 – käytetään arvioimaan pohjaveden suljetut olosuhteet, ja HCO3 – on SO42: n anaerobisen rikinpoistoreaktion tuote−, joten korkea HCO3− voidaan käyttää merkkinä hyvästä tiivistyksestä ja hiilipohjaisen pohjaveden voimakkaasta vähentämisestä . C4-alueen vesikemia on HCO3 * SO4-Ca * Mg, C7-alueen vesikemia muistuttaa C4-aluetta, HCO3 * SO4-Ca * Na, ja tarjoaa suhteellisen suljetun anaerobisen ympäristön. Tällöin vetyä tuottavien bakteerien chao1-indeksi C4: ssä on 148, Shannonin indeksi on 1,52; metanogeenien chao1-indeksi on 368, Shannonin indeksi on 2,35. Vetyä tuottavien bakteerien C7-indeksi Chao1 on 458, Shannonin indeksi on 1,98, Chao1-indeksi on 256 ja Shannonin indeksi on 2,47. C6: ssa veden kemia on SO4·HCO3–K·Na ja SO42− on dominoiva, kun taas C6: ssa vetyä tuottavien bakteerien Chao1-indeksi on 472, Shannonin indeksi on 1,56, metanogeenien Chao1-indeksi on 384 ja Shannonin indeksi on 1,08. Tiedot osoittavat, että monimuotoisuus C6 on hieman pienempi kuin C4 ja C7.
joitakin vetyä tuottavia bakteereja ja metanogeeneja havaittiin C8-ja C9-alueilla alueella, jossa biometaania ei löytynyt. Huomionarvoista oli myös, että näiden kahden alueen pohjavesiolosuhteet ovat samanlaiset kuin edellä mainituilla biogeenisillä metaanialueilla, jotka sijaitsevat pohjaveden valumavyöhykkeellä ja pohjaveden latautuminen pystyy kuljettamaan orgaanista ainesta kivihiilisaumaan, jolloin suuri määrä bakteereja kasvaa ja lisääntyy, mikä on yksi syy C8: n ja C9: n runsauteen ja monimuotoisuuteen.
lämpötila
lämpötila ja hivenaineet vaikuttavat mikrobiyhteisöjen runsauteen ja monimuotoisuuteen muuttamalla suoraan mikro-organismien kasvua ja aineenvaihduntaa sekä niiden metabolista ympäristöä. Mikrobiologiselta kannalta optimaalinen lämpötila on yksi tärkeimmistä mikro-organismien kasvuun ja aineenvaihduntaan vaikuttavista tekijöistä. Kuva 5b osoittaa, että lämpötilalla on suhteellisen heikko vaikutus metanogeenien runsauteen ja monimuotoisuuteen, vaikka vetyä tuottavien bakteerien ekologinen amplitudi on kapea ja ne ovat herkkiä lämpötilan muutoksille. Tämä muuttuja korreloi lajien runsauden ja monimuotoisuuden kanssa, ja tämän tutkimuksen tulokset osoittavat, että kivihiilisauman lämpötila (eli 25-27 °C) korreloi positiivisesti bakteerikannan runsauden kanssa. C8: ssa lämpötila oli 27,2 °C, mikä on korkein tässä tutkimuksessa mitattu lämpötila. Vetyä tuottavien bakteerien chao1-indeksi oli 510 ja Shannonin indeksi 2,61, joka on myös korkein näytteistä (C1–C8). Alin lämpötila, 24,9 °C, löytyi C1: stä, jossa bakteeriyhteisön chao1-indeksi oli alhaisin. Mikrobilajien runsaus ja monimuotoisuus lisääntyy lämpötilan ollessa C3 > C6 > C2 > C7 > C5 > C4. Geoterminen gradientti poikkeamat C9 ja C10 aiheutti paljon korkeampia lämpötiloja; Ympäristön lämpötila C9 ja C10 olivat 34.60 °C ja 40.10 °C, vastaavasti. C9: n vetyä tuottavien bakteerien Chao1-indeksi oli 176 ja Shannonin indeksi 1,28; Chao1-indeksi on 237, Shannonin indeksi 1,58. Kahdeksaan ensimmäiseen alueeseen verrattuna runsaus ja monimuotoisuus ovat hieman vähentyneet. Täällä sekä vetyä tuottavat bakteerit että metanogeenit voivat kasvaa ja lisääntyä ympäristön lämpötilassa.
mikrobien syntrofiset vuorovaikutukset
hiilisauman ääriympäristössä syntyy bakteerien yhteenliittymiä kivihiilisauman pieneliöiden kesken. Aineenvaihduntatuotteiden vaihdon ja mikroympäristössä ohjatun symbioosin kautta kilpailu ja resurssien jakaminen ylläpitävät mikrobiyhteisön erityistehtäviä, jotka määrittävät biometaanin tuotantoreitit kivihiilisaumassa. Metanotrix, joka muuttaa etikkahappoa metaaniksi, on hallitseva suku C1-alueen metanogeeniyhteisössä. Alkalibasulumiin ja Desulfosporosinukseen liittyvät bakteerit ovat homoasetogeenisia bakteereja, jotka käyttävät H2: ta elektronien luovuttajana etikkahapon tuottamiseen. Ne ovat hydrotrofisten metanogeenien pääkilpailijoita ja tarjoavat myös metanogeenien metabolista substraattia. Vetyä tuottavista bakteereista kuten Clostridiumista ja Tissierellasta saadaan myös etikkahappoa, ja siten vetyä tuottavien bakteerien runsaus tarjoaa rikkaan metabolisen substraatin Metanotrixille. Yhdessä metanogeenit ja vetyä tuottavat bakteerit ovat syntrofisessa vuorovaikutuksessa ja metaanin muodostumisreitti tällä alueella määräytyy etikkahapon hajotessa. C2: n, C4: n ja C6: n metanogeenit ovat pääasiassa hydrotrofisia metanogeeneja. Sekä hydrolyyttiset käymisbakteerit että asetogeenit edistävät etikkahapon ja H2: n tuotantoa. Ne tuottavat myös entsyymejä, kofaktoreita ja metabolisia signaaleja säätelemään vedyntuotantoa. Lisäksi homoasetogeeniset bakteerit ja asetogeenit eivät kilpaile näillä alueilla. Hydratrofiset metanogeenit voivat tuottaa metaania edellisessä vaiheessa tuotetusta CO2: sta ja H2: sta. Siksi metaboliareittejä näillä alueilla käytetään pääasiassa H2: lle, formiaatille ja muille aineille.
yli 99% C3-alueesta sisältää metyylitrofisia metanogeenejä, kuten: Metanolobus. Brevibacter, Paenibacillus, Brochothrix ja Lactococcus. Aiemmat tutkimukset osoittivat, että metoksiaromaattiset yhdisteet (tärkeä osa lignoselluloosaa) hajoavat metanolin ja muiden aineiden tuottamiseksi . Tämän alueen mikro-organismit saattavat hajottaa kivihiilen lignoselluloosan kaltaista ainesta, jolloin saadaan resursseja metyylitrofisille metanogeeneille. Tämä yksinkertainen mikrobiyhteisö ei pysty tarjoamaan riittävää substraattia Metanogeeneille, jotka kuluttavat H2: ta. Biometaanin tuotantoreitti tällä alueella perustuu metyyliyhdisteiden kulutukseen.
stafylokokkia havaittiin myös C3-alueella. Äskettäin Staphylococcus AntiMn-1 eristettiin Clarion-Clippertonin alueen syvänmeren sedimenteistä, joiden mangaanipitoisuus oli korkea. Se sisälsi geenejä, joilla oli korkea mangaaninkestävyys, minkä arvellaan olevan sopeutuma merellisiin sedimenttiympäristöihin . C3-alueen raskasmetallipitoisuus on suhteellisen korkea. Voi olla, että kivihiilisaumaympäristö voi tehokkaasti indusoida resistenssigeenien ilmentymistä, joilla voi olla antagonistisia ja detoksifioivia vaikutuksia raskasmetallien kuljetukseen ja myrkyllisyyteen mikro-organismeissa. Tämän alueen stafylokokki voi sisältää resistenssigeenejä, jotka sopeutuvat kivihiilen sauma-ympäristöön niin, että se voi osallistua myös hiilen käymisaineenvaihduntaan. C7: ssä on monia eri metanogeenilajeja, ja vetyä tuottavia bakteereja hallitsevat Clostridium, Bacillus, Citrobacter ja muut anaerobit, jotka tarjoavat substraatteja asetoklastisille metanogeeneille ja myös H2, CO2 ja formiaatti hydrotrofisille metanogeeneille. Lisäksi kertyvä etikkahappo vähentää sulfaattia pelkistäviä bakteereja, kuten Desulfosporosinusta ja Desulfitobakteereja. SRB: llä on voimakkaampi affiniteetti etikkahappoon kuin asetoklastisilla metanogeeneilla, mutta ne eivät kilpaile metyylitrofisten metanogeenien kanssa tietyistä substraateista, kuten metanolista. Siten sekä sulfaattia pelkistävien bakteerien että metanogeenien metabolismit voivat edetä tällä alueella samanaikaisesti . Metaboliaa C7: ssä hallitsivat metyylitrofiset metanogeenit, joita seurasi etikkahappokäyminen ja sitten hiilidioksidin pelkistys.