Canal KcsA: Structure
Cet article se concentrera sur KcsA, un canal potassique de Streptomyces lividans, qui contient deux domaines. Le premier étant la partie transmembranaire, qui contient 120 résidus. Cela sera discuté en détail ci-dessous. Le deuxième domaine est un domaine cytoplasmique qui contient environ 40 résidus. Le domaine cytoplasmique contribue à la stabilité de l’état fermé. Ceci est conclu sur la base de données qui montrent une diminution de l’efficacité du repliement et de l’assemblage des canaux et une incidence de la stabilité thermique lorsque le domaine cytoplasmique est retiré de la structure cristalline.5 Comme on le voit sur la figure 2, le domaine cytoplasmique C-terminal rempli d’eau est une structure de faisceau à 4 hélices qui s’étend linéairement vers le cytoplasme.5-6 Il a été démontré par les données de résonance paramagnétique électronique, EPR, produites par Uysal, que le C-terminal a un étirement qui se gonfle là où le domaine C-terminal rencontre le domaine transmembranaire.5 Ce tronçon d’hélice qui relie les deux présente un degré de flexibilité plus élevé que les autres parties qui jouent potentiellement un rôle lors de l’ouverture du canal.5 Il reste encore beaucoup de recherches à faire pour bien comprendre la fonctionnalité du domaine du terminal C. Une fonction suggérée du domaine est qu’il peut servir de récepteur pour une protéine ou un ligand activateur plasmique non identifié, mais d’autres soutiennent qu’il n’y a pas d’effet significatif sur le chemin de perméation de l’ion. La structure ne contient pas encore d’orientation de chaînes latérales, mais la disposition des hélices suggère des interactions électrostatiques dans la sous-unité.6 La figure 3 montre comment le domaine cytoplasmique se compare entre un canal ouvert et un canal fermé avec deux canaux bactériens.7 Le reste de l’article se concentrera sur le domaine transmembranaire qui sélectionne le potassium.
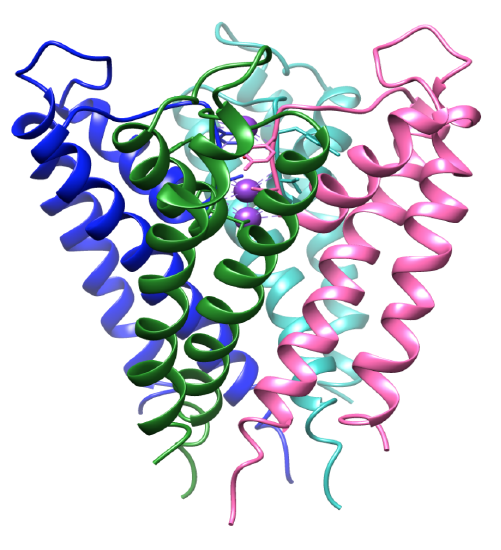
Figure 3. Le domaine transmembranaire. Chaque sous-unité est colorée différemment et les ions potassium sont violets. APB 1BL8.
La sélectivité des ions potassium dépend fortement de la structure du filtre de sélectivité des canaux potassiques. L’environnement de coordination du canal contient des ligands sélectifs qui se lient spécifiquement aux ions potassium.8 La protéine des canaux potassiques est un tétramère, qui contient quatre sous-unités identiques qui forment un pore central.8-9 Les ions s’écoulent dans un pore qui s’ouvre et se ferme, dirigeant les ions vers la cavité centrale. Les sous-unités contiennent deux hélices alpha où l’une fait face au pore central, à l’hélice interne, et l’autre à la membrane lipidique, autrement appelée hélice externe.3 Avec les hélices interne et externe, chaque sous-unité comprend une hélice de pores.3,8 Les sous-unités sont inclinées, comme on le voit sur la figure 3, où le filtre de sélectivité est près de la surface extracellulaire de la membrane. Les quatre hélices internes sont structurées de manière à se réunir près de la surface intracellulaire. Les entrées intra et extracellulaires sont chargées négativement par les acides aminés acides. Cela permet une concentration plus élevée de cations près de la membrane et moins d’anions en raison des charges opposées.8

Figure 4. Canal potassique (structure KcsA de Streptomyces lividans) avec des composants structurels clés marqués. Les principaux aspects à noter sont le filtre à pores, à cavité et à sélectivité (SF). Ce sont les parties du canal où les ions potassium traversent. Tout d’abord, l’ion s’écoule à travers le pore et pénètre dans la cavité hydratée lorsque le pore est ouvert. Ensuite, un ion potassium est déshydraté et sélectionné pour traverser le SF où il entrera ensuite dans la cellule. Notez que seules deux sous-unités sont affichées dans cette image pour une visualisation plus facile. PBD 1K4C.
La région des pores a d’abord été identifiée avec des toxines du scorpion bloquant les pores. Ils interagissent avec les acides aminés à l’entrée du pore, provoquant un blocage et un dysfonctionnement. Le pore est l’endroit où l’ion potassium entre, où il est ensuite transféré dans la cavité centrale, comme le montre la figure 4. Pendant ce temps, l’ion potassium reste hydraté.8 Le pore intracellulaire est la caractéristique du canal qui permet aux ions potassium d’entrer dans la cellule lorsqu’elle est ouverte ou d’être bloqués lorsqu’ils sont fermés. Les canaux s’ouvrent lorsqu’il y a un changement de potentiel membranaire, permettant un afflux d’ions potassium dans le filtre de la cavité. Une fois que la liaison sélective se produit dans le filtre de sélectivité, le potentiel chute à travers la membrane, fermant ainsi le pore.1 Le pore et la cavité sont revêtus hydrophobiquement, ce qui permet d’abaisser la barrière électrostatique. Il doit y avoir une barrière électrostatique plus faible afin que le coût énergétique de la déshydratation soit compensé en raison de la grande barrière énergétique pour la perméation ionique (barrière diélectrique). Ainsi, la cavité remplie d’eau permet un environnement énergétiquement favorable.3,8,10,11
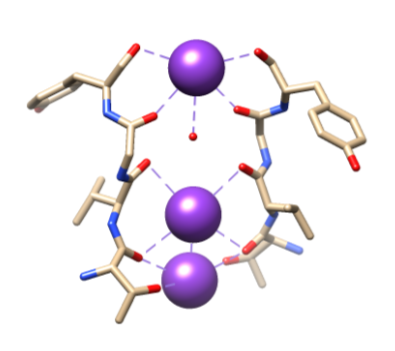
Figure 5. Liaison des ions potassium (violet) à des ligands spécifiques par les O du squelette de Thr 75, Val 76, Gly 77 et Tyr 78. La coordination complète n’est pas affichée ici pour une visualisation plus facile. Reproduit à partir de Samsom.11
Ensuite, les ions doivent être sélectionnés dans le filtre de sélectivité (SF), qui est tapissé d’atomes de chaîne principale polaire appartenant à des acides aminés. Le SF est si étroit que les ions doivent d’abord être déshydratés avant d’y pénétrer. Il y a deux caractéristiques essentielles dans le SF. Premièrement, les atomes de la chaîne principale créent des dimensions appropriées pour coordonner un ion potassium déshydraté par empilement, créant ainsi des anneaux d’oxygène séquentiels. Cela crée une très petite distance que l’ion potassium doit parcourir pour atteindre chaque site tout en traversant le filtre. Cela forme une séquence de quatre sites de coordination octuple cubiques, comme le montre la figure 6.3,8 À tout moment, ces sites sont occupés par deux ions K + et deux molécules d’eau dans une séquence alternée La deuxième caractéristique est l’emballage protéique autour du SF, qui suit une séquence de T-V-G-Y-G, illustrée à la figure 5.9 La conduction peut alors se produire lorsque deux ions potassium pénètrent dans le SF, provoquant un équilibre entre la force d’attraction qui se produit du SF à l’ion avec la force répulsive qui se produit entre les deux ions potassium.8 Cette répulsion aide les ions potassium à surmonter leur affinité intrinsèque pour se lier étroitement aux sites de liaison.3 Un examen plus approfondi du mécanisme de sélection sera discuté ci-dessous.
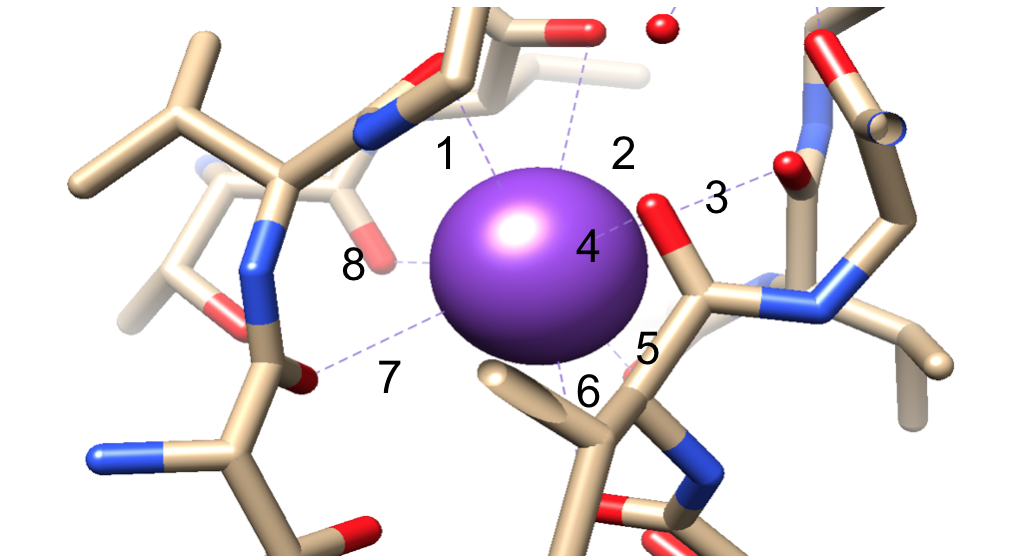
Figure 6. Un exemple d’ion potassium (violet) dans un site de coordination octuple. Reliure numérotée pour une visualisation plus facile.