KcsA Channel: Structure
Questo articolo si concentrerà su KcsA, un canale del potassio da Streptomyces lividans, che contiene due domini. Il primo è la porzione transmembrana, che contiene 120 residui. Questo sarà discusso in dettaglio di seguito. Il secondo dominio è un dominio citoplasmatico che contiene circa 40 residui. Il dominio citoplasmatico contribuisce alla stabilità dello stato chiuso. Questo è concluso sulla base di dati che mostrano una diminuzione dell’efficienza della piegatura del canale e dell’assemblaggio e la stabilità termica è influenzata quando il dominio citoplasmatico viene rimosso dalla struttura cristallina.5 Come si vede nella Figura 2, il dominio citoplasmatico C-terminale riempito d’acqua è una struttura a fascio a 4 eliche che si estende linearmente verso il citoplasma.5-6 È stato dimostrato dalla risonanza paramagnetica elettronica, EPR, dati prodotti da Uysal, che il C-terminale ha un tratto che si gonfia dove il dominio C-terminale incontra il dominio transmembrana.5 Questo tratto di elica che collega i due ha un grado di flessibilità superiore rispetto alle altre parti che potenzialmente gioca un ruolo durante l’apertura del canale.5 C’è ancora molta ricerca che deve essere condotta per comprendere appieno la funzionalità del dominio C-terminal. Una funzione suggerita del dominio è che può servire come recettore per una proteina o un ligando attivatore plasmatico non identificato, ma altri sostengono che non vi è alcun effetto significativo sul percorso di permeazione per lo ion. La struttura non contiene ancora l’orientamento di qualsiasi catena laterale, ma la disposizione delle eliche suggerisce interazioni elettrostatiche nella subunità.6 Figura 3, dimostra come il dominio citoplasmatico confronta tra un canale aperto vs un canale chiuso con due canali batteri.7 Il resto della carta si concentrerà sul dominio transmembrana che seleziona per il potassio.
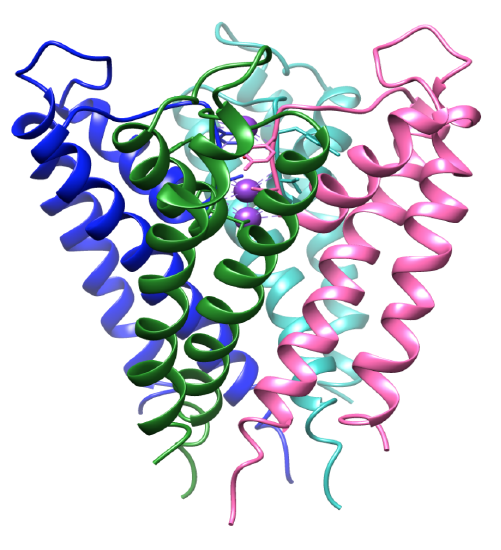
Figura 3. Il dominio transmembrana. Ogni subunità è colorata in modo diverso e gli ioni di potassio sono viola. PPB 1BL8.
La selettività per gli ioni di potassio si basa fortemente sulla struttura del filtro di selettività del canale del potassio. L’ambiente di coordinamento del canale ha ligandi selettivi che si legano specificamente agli ioni di potassio.8 La proteina del canale del potassio è un tetramero, che contiene quattro subunità identiche che formano un poro centrale.8-9 Gli ioni fluiscono in un poro che si apre e si chiude, dirigendo gli ioni verso la cavità centrale. Le subunità contengono due eliche alfa in cui si affronta il poro centrale, elica interna, e si affronta la membrana lipidica, altrimenti nota come elica esterna.3 Insieme alle eliche interne ed esterne, ogni subunità include un’elica dei pori.3,8 Le subunità sono inclinate, come si vede in Figura 3, dove il filtro di selettività è vicino alla superficie extracellulare della membrana. Le quattro eliche interne sono strutturate in un modo in cui si uniscono vicino alla superficie intracellulare. Entrambi gli ingressi intra ed extracellulari sono caricati negativamente da aminoacidi acidi. Ciò consente una maggiore concentrazione di cationi vicino alla membrana e meno anioni a causa delle cariche opposte.8

Figura 4. Canale del potassio (struttura KcsA da Streptomyces lividans) con componenti strutturali chiave etichettati. Gli aspetti principali da notare sono il poro, la cavità e il filtro di selettività (SF). Queste sono le parti del canale in cui viaggiano gli ioni di potassio. In primo luogo, lo ion scorre attraverso il poro ed entra nella cavità idratata quando il poro è aperto. Quindi uno potassium di potassio viene disidratato e selezionato per viaggiare attraverso la SF dove entrerà nella cella. Si noti che solo due subunità sono mostrate in questa immagine per facilitare la visualizzazione. PBD 1K4C.
La regione dei pori è stata identificata per la prima volta con le tossine dello scorpione che bloccano i pori. Interagiscono con gli amminoacidi all’ingresso del poro causando blocco e disfunzione. Il poro è dove entra lo potassium potassio, dove poi viene trasferito alla cavità centrale, come si vede in Figura 4. Durante questo periodo lo ion di potassio rimane idratato.8 Il poro intracellulare è la caratteristica del canale che consente agli ioni di potassio di entrare nella cellula quando è aperta o di essere bloccati dall’ingresso quando è chiuso. I canali si aprono quando c’è un cambiamento nel potenziale di membrana, consentendo un afflusso di ioni di potassio nel filtro della cavità. Una volta che il legame selettivo si verifica nel filtro di selettività, il potenziale scende attraverso la membrana, chiudendo così il poro.1 Il poro e la cavità sono idrofobicamente allineati, permettendo la capacità di abbassare la barriera elettrostatica. Ci deve essere una barriera elettrostatica più bassa in modo dal costo energetico di disidratazione è compensato dovuto la grande barriera di energia per la permeazione dello ion (barriera dielettrica). Pertanto, la cavità riempita d’acqua consente un ambiente energeticamente favorevole.3,8,10,11
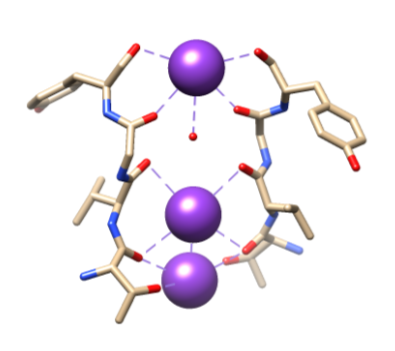
Figura 5. Legatura degli ioni del potassio (porpora) ai leganti specifici dalle O della spina dorsale di Thr 75, di Val 76, di Gly 77 e di Tyr 78. Il coordinamento completo non è mostrato qui per facilitare la visualizzazione. Riprodotto da Samsom.11
Successivamente, gli ioni devono essere selezionati nel filtro di selettività (SF), che è rivestito con atomi di catena principale polare che appartengono agli amminoacidi. La SF è così stretta che gli ioni devono prima essere disidratati prima di entrarvi. Ci sono due caratteristiche essenziali nella SF. Innanzitutto, gli atomi della catena principale creano dimensioni adeguate per coordinare uno potassium di potassio disidratato impilando, rendendo così anelli di ossigeno sequenziali. Questo crea una distanza molto piccola che lo potassium potassio deve viaggiare per raggiungere ogni sito durante il viaggio attraverso il filtro. Questo forma una sequenza di quattro cubici siti di coordinamento otto volte, come si vede in Figura 6.3,8 In qualsiasi momento, questi siti sono occupati da due ioni K+ e due molecole di acqua in un’alternanza la seconda caratteristica è La proteina di imballaggio intorno alla SF, che segue una sequenza di T-V-G-Y-G, mostrato in Figura 5.9 Conduzione può quindi verificarsi quando due ioni potassio inserire la SF, causando un equilibrio tra la forza di attrazione che si verifica da SF a ioni di litio con la forza di repulsione che si verifica tra due ioni potassio.8 Questa repulsione aiuta gli ioni di potassio a superare la sua affinità intrinseca per il legame strettamente ai siti di legame.3 Ulteriori discussioni sul meccanismo di selezione saranno discusse di seguito.
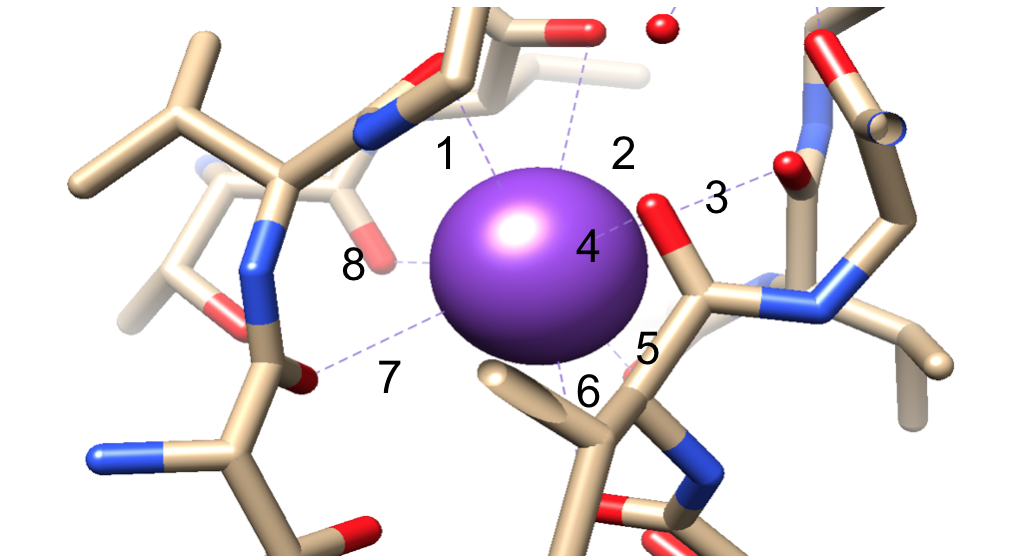
Figura 6. Un esempio di uno potassium di potassio (viola) in un sito di coordinazione ottuplice. Rilegatura numerata per una visualizzazione più semplice.