微生物群集構造に影響を与える要因の分析
環境要因とコミュニティ分散との関係をさらに調べるために、RDA分析を作成しました。 7). 合計で、微量元素Fe、Co、Ni、温度、塩分、深さ、水分、およびROを含む八つの環境要因、。 採取場所での深さと貯留層温度を測定し、その他の情報は地元の鉱山の地質データから得られた(表2参照)。 データを図1 0に示す。 7aは、この研究で見出された細菌群集組成が、Fe、Ni、水分、塩分、およびROによって有意に影響されることを明らかにした。 C4、C7、C10以外のすべてのコミュニティはROと正の相関があり、C4、C7、およびC10はFe、Ni、および水分と正の相関があります。 Coは、メタン生成物質の生化学的代謝において重要な酵素である補酵素Mメチルトランスフェラーゼに必要である ; したがって,古細菌群集に対するCoの効果は細菌群集のそれよりも大きい。 F e,Co,Niの元素は水分と同様に,古細菌群ではR oと塩分に続いて最も重要な環境要因であると思われた。 CoとC1、C8、C9、C10コミュニティとの間には有意な正の相関があります。 C2、C4、C6、およびC5を除くすべてのコミュニティは、塩分と負の相関を示した(図。 7b)。

石炭床環境要因と石炭特性を持つ細菌(a)と古細菌(b)のレベルに基づくrda(冗長性分析)。 衝撃係数の長さは長く、衝撃の寄与は高く、逆に衝撃係数が短い場合、衝撃の寄与はより軽い。 環境要因がサンプルと鋭く曲がるとき、正の相関関係があり、環境要因およびサンプル角度が鈍角のとき、否定的な相関関係があります
石炭ランク
石炭化”ジャンプ”とは、地質学的歴史の中で石炭の温度と圧力の下で一連の物理的および化学的変化を指します。 石炭は、このように突然の変化に緩やかなプロセスを経ています。 四つのジャンプは0.6、1のROに対応しています。3、2.5、および3.0%。 検討中の古細菌や細菌群集にかかわらず、石炭ランクは細菌の多様性と豊富さに一定の影響を与えます。 石炭ランクの上昇に伴い、古細菌群集と細菌群集の両方で、コミュニティの多様性は全体的に一定の減少傾向を示している(図。 8). さらに、微生物は、石炭ランクによって制御される石炭の組成に影響を与える可能性がある。 中-低ランクの石炭は、植物進化物質を多く含むグループ1の植物進化物質を大量に含有しています。 ここでは、水素、酸素、窒素の含有量が高く、細菌が必要とする栄養素が豊富です。 石炭化の過程で、有機物は多くの水分と液体炭化水素を生成します。 同時に、石炭に含まれる水素と酸素の側鎖も豊富である。 これらの液体および固体物質は、細菌の生命の基盤を提供する。 その結果、この地域の石炭中の水素産生細菌およびメタン生成物質の豊富さおよび多様性は比較的高い。 Roの増加に伴い、石炭中の水素および酸素の側鎖含有量が大幅に減少し、微生物に利用可能な成分も減少する。 したがって、グループ2およびグループ3の細菌および古細菌コミュニティの種の豊富さおよび多様性は、全体的に減少する。 これまでのところ、生物起源の炭層メタンの石炭ランクは、2.0%(C4Hebi)の反射率を有することが自然界で発見されている。 RO>2.5%の後、小分子に変換できる有機化合物は非常にまれですが、グループ4では多様性と豊富さが高くなっています。 この時点で地下水によって導入された栄養素は、細菌の再生に利用可能であると推測している。 異なる地域と異なる季節の地下水によってもたらされる栄養成分は、種の多様性に寄与している可能性がある。 グループ4の植物相の多様性が高い理由の一つは、C8焦作九栗山エリアがより良い地下水流出条件とより強い再充電を持っていることがあります。 それは植物相のための栄養素を輸送することができるので、多様性と豊富さはグループ2よりも高いです。 古細菌群集の多様性と豊富さは、石炭ランクとある程度負の相関があることは注目に値する。 しかし,細菌群集における種の豊富さは石炭ランクと正の相関を示し,多様性は減少傾向を示した。 石炭ランクの上昇に伴い、いくつかの細菌群は徐々に様々な石炭ランクの環境に適応し、成長し、大量に増殖することができ、メタン生成物質は石炭ランクに適応することは困難である。
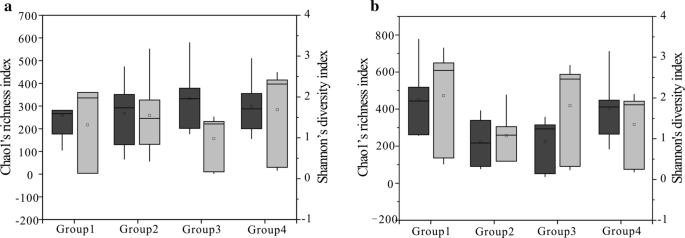
Chao1(ダークグレー)とシャノン(ライトグレー)の四つのグループのインデックス(石炭サンプルは、細菌コミュニティ(a)と古細菌コミュニティ(b)のROの値に応じて四つのグループに分けられ、1グループは0.6%未満、2グループは0.8と1.1%の間の値、3グループは1.4と1.8%の間の値、4グループは2.67と3.15%の間の値を表している)地域由来の値。 25番目と75番目の百分位数はボックスの外側の端で示され、最大値と最小値はウィスカーの端で示され、中央値は各ボックス内の水平線で示されます
微量金属元素
微量金属元素は、細胞が代謝調節を通じて元素の恒常性を維持する一定の範囲内で微生物の増殖を促進することができます。 微量金属元素は様々な酵素にも存在し、嫌気性代謝の過程で微生物によって吸収され使用され、水素産生細菌およびメタン生成物質の共同体構造に影
FeとNiはCoよりも水素生成細菌に大きな影響を与えます。 FeおよびNiは、微生物中のヒドロゲナーゼおよび他の金属酵素の合成および代謝に関与することができる。 FeおよびNiの含有量が一定の範囲内で増加するにつれて、水素産生細菌集団の豊富さおよび多様性も増加する。 C7中のFeとNiの含有量は他の地域よりもはるかに高く、この研究はClostridiumが水素を産生する細菌であることを発見しました。 この知見は、微量元素の過剰なレベルが微生物の増殖に毒性作用を有し、金属酵素の活性を阻害する可能性があることを示している。 C4、C6、およびC9中のFeのレベルは有意に異ならず、3500mg Kg−1で安定であった(図10B)。 9). 3つの領域におけるNiの相対的な存在量はc6>C9>C4であり、これは存在量の順序(c6>C9>C4)に対応するが、多様性の順序はC6>C4>C9である。 属のメンバー:Clostridium、Klebsiella、EnterobacterとCitrobacterは、C4、C6およびC9コミュニティで検出された;他の地域よりも高い存在量と多様性のレベルでそれらを含む。

石炭サンプル中のFe、Co、Niの微量金属元素含有量
古細菌群集では,メタン生成物質に対するFe,Co,Niの影響はさらに重要である。 Coはメタン生成補酵素F430の合成における重要な要素であり、上位3つのCo含有量はC8>C7>C10であり、メタン生成種および豊富さはC8>C7>C10である。 Coの含有量は,メタノゲンの豊富さと多様性とある程度正の相関を示した。 C7中のFeの含有量は他の領域のそれよりもはるかに高いが、それは領域内のメタン生成物質の分布に影響を与えません。 これまでの研究では、Methanothermobacterと命名された唯一の種が発見されました。 M.marburgensisのメタノーゲンにおけるモノフェリックヒドロゲナーゼの存在は、メテニル-H4MPT+とh2の可逆反応を触媒し、メチレン-H4MPTとH+を生成し、CO2とh2からメタンを生成する。 水素栄養代謝を用いたメタン生成物にも同様の酵素が含まれている可能性がある。 さらに、methanoculleusとMethanobacteriaを除いて、methanotrophic methanogensの大部分はこのような酵素を保有していると推測されています。 Methylotrophic methanogenのFeと関連付けられるmetalloenzymeがあるかもしれないことが推測されます。
地下水の条件
地下水は、直接的または間接的に、炭層における極端な生物の成長と代謝のための生態学的基盤を提供する。 一方で、地下水の再充電は、細菌や古細菌のコミュニティのための栄養素を大量に供給します; 一方、地下水環境条件(Eh、pH、塩分、イオン組成、および微量元素)は、微生物の増殖および代謝酵素活性に直接影響する。 地下水環境条件は石炭の使用と劣化に直接関係しており、炭層に位置する微生物は異なるコミュニティ構造と機能特性を示しています。
鉱業地域の流出ゾーンは、石炭の継ぎ目の生存を可能にすることができます。 高透過性貯留層は水素産生細菌およびメタン生成物質の増殖および再生にプラスの影響を与えるが、変成作用は石炭貯留層の石炭透過性に重大な負の影響を及ぼす。 生物起源のCBMがある地域では、C2、C4、C6、およびC7コミュニティはすべて十分に文書化されています。 これらのコミュニティは、低および中程度の石炭ランクに属し、石炭の気孔率は高ランクの石炭よりも比較的高く、地下水は時間内に炭層の微生物に養分を提供することができます。 米国のパウダー川流域内の現在のCBM開発ゾーン 主に地下水流出ゾーンに集中しています。 C6採掘地域の浅いCBM井戸からのガス安定同位体データはまた、地域における生物遺伝的CBMの存在を確認した。 しかし,別の深いCBM井戸からのガス安定同位体データは,CBMが主に熱発生性であることを示した。 これらの結果は、埋葬の深さが増加するにつれて、流出条件が弱まり、微生物のための栄養素を輸送することが困難になり、コミュニティの豊富さと多様性が低下することを示している。 屋根と床のNo. 2C2地域の炭層は泥岩と粘土岩の比較的安定した層を有しており、炭層内の水素産生細菌やメタン生成菌が液体栄養素を得ることが困難であり、その成長と代謝を制限し、したがってコミュニティの多様性と豊富さを制限している。 この領域では、Chao1指数は240であり、シャノン指数は細菌群集で1.38であることに注意してください。 メタノゲンのChao1指数は82であり、シャノン指数は0.56である。 砂岩の裂けた帯水層の屋根は、Noのc4領域にあります。 21炭層は、より良い充電条件を持っており、水で炭層を満たしています。 微生物群集は、輸送に影響を与える異なる栄養素の十分な利用可能性から累積的な効果を経験する可能性があり、C2領域の微生物群集の豊富さと多様性と比較して、大幅に改善されている。 この領域では、水素産生細菌のChao1指数は148、シャノン指数は1.52、メタン生成菌のChao1指数は368、シャノン指数は2.35である。 C6区域の砂岩の割れ目帯水層は第3炭層の直接水で満たされた帯水層である。 層内の破壊発達と中程度の水圏はまた、コミュニティの豊かさと多様性に積極的な役割を果たしています。 ここでは、水素産生細菌のChao1指数は472、シャノン指数は1.56、メタン生成菌のChao1指数は384、シャノン指数は1.08である。 これはまた、C7地域の場合であり、第5炭層は良好な再充電条件を有する直接破砕帯水層を有し、水素産生細菌の存在度指数は458であり、シャノン指数は1.98である。; メタノゲンのChao1指数は256であり、シャノン指数は2.47である。 したがって、c4、C6、およびC7における水素産生細菌およびメタン生成物質の種の多様性は、C2のそれよりも高い。
地下水の環境条件は、微生物の成長と代謝に直接影響します。 石炭床地下水のpH値は一般的に中性であるが、pH値は6.5から8.4の間で変化し、c4エリアNo.21炭層の砂岩破砕帯水層では塩分は1.0g L-1である。 Noの直接帯水層では。 3炭層C6エリアでは、pHはpH6.8から8.0の範囲であり、塩分は0.7g L−1である。 C7領域の地下水pH値は6.1–7.3であり、塩分は1.25g L−1である。 C4、C6、およびC7のpH値は中性に近く、鉱化の程度は低く、微生物群集はより良好な成長、より高い豊富さおよびより高い多様性を有する。 さらに,地下塩濃度とイオン組成は炭層の嫌気性還元環境と密接に関連している。 例えば、SO42−は地下水の閉鎖状態を評価するために使用され、HCO3−はSO42−の嫌気性脱硫反応の産物であり、高いHCO3-は良好な密閉性と石炭床地下水の強 C4領域の水化学はHCO3·SO4–Ca·Mgであり、C7領域の水化学はC4領域HCO3·SO4–Ca·Naと同様であり、比較的閉鎖された嫌気性環境を提供する。 この場合、C4中の水素産生細菌のChao1指数は148であり、Shannon指数は1.52である; メタノゲンのChao1指数は368であり、シャノン指数は2.35である。 C7中の水素産生細菌のChao1指数は458、シャノン指数は1.98、Chao1指数は256、シャノン指数は2.47である。 C6では、水化学はSO4·HCO3–K·Naであり、SO42−が支配的であるが、c6の水素生産細菌のChao1指数は472であり、シャノン指数は1.56であり、メタン生成物質のChao1指数は384であり、シャノン指数は1.08である。 データは、C6の多様性がC4およびC7よりもわずかに低いことを示しています。
バイオメタンが検出されなかった地域のC8およびC9領域では、水素産生細菌およびメタン生成菌が検出されました。 また、これら二つの地域の地下水の条件は、地下水流出ゾーンに位置する上記の生物起源メタン地域のものと同様であり、地下水の再充電は、石炭層にいくつかの有機物を輸送することができ、多数の細菌が増殖し、増殖することが注目され、これがC8およびC9の種の豊富さおよび多様性の高い理由の一つである。
温度
温度と微量金属元素は、微生物の成長と代謝とその代謝環境の両方を直接変化させることによって、微生物群集の存在量と多様性に影響 したがって、微生物学的観点から、最適温度は、微生物の増殖および代謝に影響を与える最も重要な要因の一つである。 図5bは、水素産生細菌が狭い生態学的振幅内に存在し、温度変化に敏感であっても、温度がメタン生成物質の存在量と多様性に比較的弱い影響を及ぼすことを示している。 この変数は種の豊富さと多様性と相関しており、本研究の結果は、石炭層の温度(すなわち、25と27℃の間)が細菌集団の豊富さと正の相関があることを示 C8では、温度は27.2℃であり、この研究で記録された最高温度であった。 水素産生細菌のChao1指数は510であり、シャノン指数は2.61であり、サンプル(C1-C8)の中でも最も高かった。 最低温度、24.9°Cは、細菌のコミュニティのChao1指数が最も低かったC1で発見されました。 微生物種の豊富さと多様性は、c3>C6>C2>C7>C5>C4の温度とともに増加します。 C9とC10の地熱勾配異常ははるかに高い温度を引き起こし、C9とC10の周囲温度はそれぞれ34.60℃、40.10℃であった。 C9における水素産生細菌のChao1指数は176であり、シャノン指数は1.28であり、Chao1指数は237であり、シャノン指数は1.58である。 最初の8つの地域と比較して、豊かさと多様性はわずかに減少しています。 ここでは、水素産生細菌とメタン生成菌の両方が周囲温度で増殖して再現することができます。
微生物相相互作用
石炭層の極端な環境では、石炭層の微生物の間に細菌のコンソーシアムが形成される。 代謝産物の交換とミクロ環境制御された共生を通じて、競争と資源配分は、炭層におけるバイオメタン生産経路を決定する微生物群集の特定の機能を維持する。 酢酸をメタンに変換するMethanothrixは、C1地域のmethanogenコミュニティの支配的な属です。 AlkalibaculumおよびDesulfosporosinusと関連付けられる細菌は酢酸を作り出すのに電子提供者としてH2を使用するhomoacetogenic細菌です。 それらはhydrogenotrophic methanogensのための主要な競争相手で、またmethanogensに新陳代謝の基質を提供します。 ClostridiumやTissierellaなどの水素産生細菌も酢酸を提供するため、水素産生細菌の豊富さはMethanothrixの豊富な代謝基質を提供します。 一緒に、メタン生成物質と水素産生細菌はsyntrophic相互作用しており、この領域におけるメタン生成経路は酢酸の分解によって決定される。 C2、C4、およびC6のメタン生成物質は、主に水素化栄養性メタン生成物質である。 加水分解発酵細菌およびアセトゲンの両方が酢酸およびH2の産生に寄与する。 それらはまた、水素産生を調節するための酵素、補因子および代謝シグナルを産生する。 なお、homoacetogenic細菌およびacetogenはこれらの区域で競いません。 水素栄養性メタン生成物質は、前の段階で生成されたCO2とH2からメタンを生成することができます。 従って、これらの区域の新陳代謝の細道はH2、ギ酸塩および他の物質のために主に使用されます。
C3領域の99%以上がMethanolobusのようなmethanotrophic methanogenを保有しています。 Brevibacter、Paenibacillus、Brochothrix、およびLactococcus。 これまでの研究では、メトキシ芳香族化合物(リグノセルロースの重要な部分)が分解されてメタノールなどの物質が生成することが示されていました。 この地域の微生物は、メチル栄養性メタン生成物質に資源を提供するために石炭のリグノセルロース様物質を分解する可能性がある。 この単純な微生物群集は、H2を消費するメタン生成物質のための十分な基質を提供することはできません。 この分野でのバイオメタン生産経路は、メチル化合物の消費に基づいています。
C3領域でもブドウ球菌が検出された。 最近、Staphylococcus AntiMn-1は、高いマンガン含有量を有するClarion-Clipperton地域の深海sed積物から単離された。 これは、海洋堆積環境への適応であると考えられているマンガンに対する高い耐性を有する遺伝子を含んでいた。 C3区域の重金属の内容は比較的高いです。 炭層環境は耐性遺伝子の発現を効果的に誘導することができ,微生物内の重金属の輸送および毒性に拮抗的および解毒効果を有する可能性がある。 この地域のブドウ球菌には、石炭の発酵代謝にも関与するように、炭層環境に適応する耐性遺伝子が含まれている可能性があります。 C7には多くの異なる種のメタン生成菌があり、水素生成細菌はクロストリジウム、バチルス、シトロバクター、および他の嫌気性菌によって支配されており、アセトクラスティックメタン生成菌の基質を提供し、水素栄養性メタン生成菌のためのH2、CO2、およびギ酸塩も提供している。 さらに、酢酸を蓄積すると、DesulfosporosinusやDesulfitobacteriumなどの硫酸還元細菌が減少します。 SRBは酢酸に対してアセトクラスティックメタノゲンよりも強い親和性を有するが、メタノールのような特定の基質についてはメチル栄養性メタノゲンと競合しない。 したがって、硫酸還元細菌とメタン生成菌の両方の代謝は、この領域で同時に進行することができる。 C7の代謝は、酢酸発酵し、二酸化炭素の減少に続いて、メチル栄養性メタン生成によって支配された。