analyse van factoren die de microbiële gemeenschapsstructuur beïnvloeden
om de mogelijke relaties tussen de milieufactoren en variantie in de Gemeenschap verder te onderzoeken, werd een RDA-analyse gemaakt( Fig. 7). In totaal acht omgevingsfactoren, waaronder sporenelementen Fe, Co, Ni, temperatuur, zoutgehalte, diepte, vocht en RO. De diepte en de temperatuur van het reservoir werden gemeten op de plaats waar de monsters werden genomen en andere gegevens werden verkregen uit de geologische gegevens van de plaatselijke mijnen (zie Tabel 2). Gegevens weergegeven in Fig. 7a bleek dat de bacteriële gemeenschap samenstellingen gevonden in deze studie werden aanzienlijk beïnvloed door Fe, Ni, vocht, zoutgehalte, en RO. Alle gemeenschappen, behalve C4, C7, C10, zijn positief gecorreleerd met RO; C4, C7 en C10 positief gecorreleerd met Fe, Ni en vocht. Co is vereist voor co-enzym m methyl-transferase, een belangrijk enzym in het biochemische metabolisme van methanogenen ; daarom is het effect van Co op de archaeale gemeenschap groter dan dat van bacteriële gemeenschap. De elementen Fe, Co en Ni, evenals vocht, bleken de belangrijkste omgevingsfactoren te zijn, gevolgd door RO en zoutgehalte in de archaeal gemeenschap. Er is een significante positieve correlatie tussen Co en de C1 -, C8 -, C9-en C10-gemeenschappen. Alle gemeenschappen behalve C2, C4, C6 en C5 waren negatief gecorreleerd met zoutgehalte (Fig. 7 ter).
Fig. 7
de RDA (redundancy analysis) gebaseerd op het niveau van bacteriën (a) en archaea (b) met de omgevingsfactoren van het kolenbed en de kolenkarakteristieken. De lengte van de impactfactor is langer, de bijdrage van het effect is hoger, en omgekeerd, wanneer de impactfactor korter is, is de bijdrage van het effect lichter. Wanneer de omgevingsfactor acuut met de steekproef wordt gehoekt, is er een positieve correlatie, en wanneer de omgevingsfactor en de steekproefhoek stomp zijn, is er een negatieve correlatie
Tabel 2 milieuinformatie en kolensteekproeven
de “sprong” verwijst naar een reeks fysische en chemische veranderingen onder de temperatuur en druk van steenkool tijdens de geologische geschiedenis. De kolen hebben aldus een proces van geleidelijke tot plotselinge verandering ondergaan. De vier sprongen komen overeen met RO van 0,6, 1.3, 2,5 en 3,0%. Ongeacht de archaeal of de bacteriële gemeenschap in kwestie, de kolen rang heeft een zekere invloed op de diversiteit en de overvloed van de bacteriën. Met een toename van de rang van steenkool, zowel in de archaeale als in de bacteriële gemeenschappen, vertoont de diversiteit van de Gemeenschap een zekere neerwaartse trend (Fig. 8). Bovendien kunnen micro-organismen van invloed zijn op de samenstelling van de door de kolenlagen gecontroleerde steenkool. De middelste en lage kolen bevatten grote hoeveelheden plant geëvolueerde stoffen in Groep 1, die veel plant geëvolueerde stoffen bevat. Hier is er een hoger gehalte aan waterstof, zuurstof en stikstof; en de voedingsstoffen die de bacteriën nodig hebben zijn overvloedig. Tijdens het proces van coalificatie genereren organische stoffen veel vocht en vloeibare koolwaterstoffen. Tegelijkertijd zijn ook de zijketens van waterstof en zuurstof in steenkool overvloedig aanwezig. Deze vloeibare en vaste stoffen vormen de basis van het leven voor bacteriën. Als gevolg daarvan zijn de overvloed en diversiteit van waterstofproducerende bacteriën en methanogenen in steenkool in deze regio relatief hoog. Met de toename van Ro wordt het zijketengehalte van waterstof en zuurstof in steenkool drastisch verminderd en worden ook de voor de micro-organismen beschikbare componenten verminderd. Daarom zijn de soortendichtheid en diversiteit van de bacteriële en archaeale gemeenschappen in groep 2 en groep 3 in het algemeen verminderd. Tot nu toe is gebleken dat de steenkoollagen van biogeen koolbed methaan in de natuur een reflectiviteit hebben van 2,0% (C4 Hebi). Na RO > 2,5% zijn de organische verbindingen die in kleine moleculen kunnen worden omgezet zeer zeldzaam geweest, maar er is een grotere diversiteit en overvloed in groep 4. We speculeren dat de voedingsstoffen die op dit moment door grondwater worden geïntroduceerd, beschikbaar zijn voor bacteriële voortplanting. De nutriënten die grondwater in verschillende regio ‘ s en in verschillende seizoenen teweegbrengt, kunnen hebben bijgedragen aan de diversiteit van soorten. Een reden voor de grotere flora diversiteit in groep 4 kan zijn dat C8 Jiaozuo Jiulishan gebied heeft betere grondwaterafvoer omstandigheden en sterkere aanvulling. Het kan voedingsstoffen transporteren voor de flora, dus de diversiteit en overvloed zijn groter dan Groep 2. Het is vermeldenswaard dat de diversiteit en de overvloed van de archaeal gemeenschappen zijn negatief gecorreleerd met kolen rangen tot op zekere hoogte. Echter, de soortenrijkdom in de bacteriële gemeenschappen is positief gecorreleerd met kolen rangen, en de diversiteit toont een neerwaartse trend. Met de opkomst van kolenlagen, sommige bacteriële groepen geleidelijk aangepast aan de omgeving van verschillende kolenlagen en kunnen groeien en vermenigvuldigen in grote aantallen, en methanogenen zijn moeilijk aan te passen aan kolenlagen.
Fig. 8
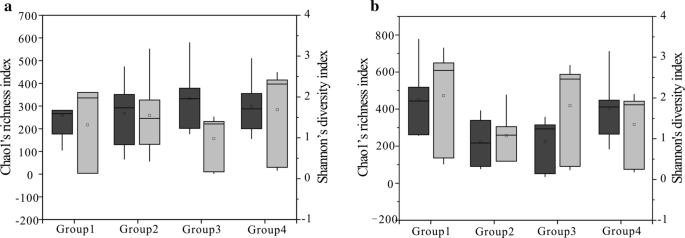
Chao1 ’s (donkergrijs) en Shannon’ s (lichtgrijs) index voor de vier groepen (Kolenmonsters werden in vier groepen verdeeld volgens de waarde van RO voor de bacteriële gemeenschap (A) en archaea gemeenschap (b), groep 1 vertegenwoordigt een waarde van minder dan 0,6%, groep 2 vertegenwoordigt de waarde tussen 0,8 en 1,1%, groep 3 vertegenwoordigt de waarde tussen 1,4 en 1,8%, groep 4 vertegenwoordigt de waarde tussen 2,67 en 3,15%) afgeleid van regio ‘ s. 25e en 75e percentiel worden aangegeven door de buitenranden van de dozen, terwijl de maximum – en minimumwaarden worden aangegeven door de uiteinden van de snorharen en de mediaan door een horizontale lijn binnen elke doos
Sporenmetaalelementen
sporenmetaalelementen kunnen de groei van micro-organismen binnen een bepaald bereik bevorderen, waar de cel de homeostase van de elementen handhaaft door metabole regulatie. Metalen sporenelementen kunnen ook voorkomen in verschillende enzymen, die door micro-organismen kunnen worden geabsorbeerd en gebruikt in het proces van anaërobe stofwisseling, wat van invloed is op de gemeenschapsstructuur van waterstofproducerende bacteriën en methanogenen (Tabel 3).
Tabel 3 de rol van Fe, Co en Ni in reactie en transformatie in anaeroob metabolisme
Fe en Ni hebben een groter effect op waterstofproducerende bacteriën dan Co . Fe en Ni kunnen deelnemen aan de synthese en het metabolisme van hydrogenasen en andere metalloenzymes in micro-organismen. Naarmate het gehalte aan Fe en Ni binnen een bepaald bereik toeneemt, neemt ook de overvloed en diversiteit van waterstofproducerende bacteriële populaties toe. Het gehalte aan Fe en Ni in C7 is veel hoger dan in andere regio ‘ s, en uit dit onderzoek is gebleken dat Clostridium waterstofproducerende bacteriën zijn. Deze bevinding wijst erop dat bovenmatige niveaus van de sporenelementen een toxisch effect op de groei van micro-organismen kunnen hebben en de activiteit van metalloenzymes kunnen remmen. De niveaus van Fe in C4, C6 en C9 waren niet significant verschillend en waren stabiel op 3500 mg Kg-1 (Fig. 9). De relatieve abundantie van Ni in de drie gebieden is C6 > C9 > C4, wat overeenkomt met de abundantieorde (ook C6 > C9 > C4), maar de volgorde van diversiteit is C6 > C4 > C9. Leden van de geslachten: Clostridium, Klebsiella, Enterobacter en Citrobacter werden aangetroffen in de C4 -, C6-en C9-gemeenschappen, waaronder die met hogere abundanties en niveaus van diversiteit dan in andere regio ‘ s.
Fig. 9
het gehalte aan sporenmetaalelementen van Fe, Co, Ni in koolmonsters
in de archaeal gemeenschap is de invloed van Fe, Co en Ni op de methanogenen nog belangrijker. Co is een belangrijk element in de synthese van methanogeen co-enzym F430 , en het Co-gehalte in de top drie is C8 > C7 > C10, waarbij methanogene soorten en abundantie C8 > C7 > C10 zijn. Het gehalte aan Co is positief gecorreleerd met de overvloed en diversiteit van methanogenen tot op zekere hoogte. Hoewel het gehalte aan Fe in C7 veel hoger is dan dat in andere regio ‘ s, heeft het geen invloed op de distributie van methanogenen in de regio. Er zijn maar weinig soorten methanogenen die Fe kunnen bevatten – in eerdere studies werd slechts één soort, genaamd Methanothermobacter, ontdekt. De aanwezigheid van monoferrische hydrogenase in methanogenen van M. marburgensis katalyseert de reversibele reactie van methenyl-H4MPT+ en H2 om methyleen-H4MPT en H+ te produceren; het produceren van methaan uit CO2 en H2 . Methanogenen die hydrogeen metabolisme gebruiken, kunnen ook soortgelijke enzymen bevatten. Daarnaast wordt ook gespeculeerd dat een groot deel van methylotrofe methanogenen dergelijke enzymen bevat, behalve de Methanoculleus en Methanobacteriën. Er wordt gespeculeerd dat er een Metalloenzym verbonden aan Fe in de methylotrophic methanogens kan zijn.
grondwatercondities
grondwater vormt direct of indirect een ecologische basis voor de groei en het metabolisme van extremofielen in de kolenlaag. Enerzijds levert grondwateraanvulling grote hoeveelheden voedingsstoffen voor bacteriële en archaeale gemeenschappen; aan de andere kant beïnvloeden de omgevingsomstandigheden van het grondwater (Eh, pH, zoutgehalte, ionensamenstelling en sporenelementen) direct de microbiële groei en metabole enzymactiviteit. De milieuomstandigheden van het grondwater houden rechtstreeks verband met het gebruik en de afbraak van steenkool, en de micro-organismen in de kolenlaag vertonen verschillende gemeenschapsstructuren en functionele kenmerken.
microbiële nutriëntsubstraten zijn over het algemeen opgelost. De afvoerzone in het mijngebied kan het overleven van kolenlagen mogelijk maken. Reservoirs met hoge permeabiliteit hebben een positief effect op de groei en reproductie van waterstofproducerende bacteriën en methanogenen, terwijl metamorfisme een significant negatief effect heeft op de permeabiliteit van kolenreservoirs . In gebieden met biogene CBM zijn de C2 -, C4 -, C6-en C7-gemeenschappen goed gedocumenteerd. Deze gemeenschappen behoren tot de lage en middelgrote kolen rang, de porositeit van steenkool is relatief hoger dan high rank steenkool, grondwater kan voedingsstoffen leveren aan de microben in de kolenlaag in de tijd. De huidige CBM-ontwikkelingszone binnen het Powder River Basin in de VS is voornamelijk geconcentreerd in de grondwaterafvoerzone. De gasstabiele isotoopgegevens van een ondiepe CBM-put in het C6-mijngebied bevestigden ook de aanwezigheid van biogenetische CBM in het gebied. Gasstabiele isotoopgegevens van een andere diepe CBM gaven echter goed aan dat de CBM voornamelijk thermogeen is. Deze resultaten tonen aan dat naarmate de diepte van de begraving toeneemt, de afvoervoorwaarden zullen verzwakken en het moeilijk zal zijn om voedingsstoffen voor het micro-organisme te transporteren, wat zal resulteren in een afname van de overvloed en diversiteit van de gemeenschap. Het dak en de vloer van de No. 2 kolenlagen in C2-gebied hebben relatief stabiele lagen van mudstone en kleigesteente, wat het moeilijk maakt voor de waterstofproducerende bacteriën en methanogenen in de kolenlaag om vloeibare nutriënten te verkrijgen, en hun groei en metabolisme en, dus, hun gemeenschap diversiteit en overvloed beperkt. Merk op dat op dit gebied de Chao1-index 240 is en de Shannon-index 1,38 in de bacteriegemeenschap. De Chao1-index van methanogenen is 82, de Shannon-index is 0.56. De zandsteen gespleten aquifer dak in C4 gebied van de No. 21 kolenlaag heeft betere laadomstandigheden en vult de kolenlaag met water. Het is mogelijk dat de microbiële gemeenschap cumulatieve gevolgen van de voldoende beschikbaarheid van verschillende nutriënten ervaart, die vervoer beà nvloedt, vergeleken met de overvloed en de diversiteit van de micro-organismen gemeenschap in het C2-gebied, dat zeer heeft verbeterd. Op dit gebied is de Chao1-index van waterstof-producerende bacteriën 148, De Shannon-index is 1,52; de Chao1-index van methanogenen is 368, en de Shannon-index is 2,35. De zandsteen gespleten aquifer van C6 gebied is een directe water-gevulde aquifer van No. 3 kolenlaag. Breukontwikkeling binnen de laag en matige aqueositeit speelt ook een actieve rol in de overvloed en diversiteit van de gemeenschap. Hier is de Chao1-index van waterstof-producerende bacteriën 472, de Shannon-index is 1,56; de Chao1-index van methanogenen is 384 en de Shannon-index is 1,08. Dit is ook het geval in het C7-gebied, de No. 5-kolenlaag heeft een direct gebroken aquifer met goede oplaadomstandigheden, de abundantie-index van de waterstofproducerende bacteriën is 458, de Shannon-index is 1.98; en de Chao1-index van de methanogenen is 256 en de Shannon-index is 2,47. Daarom is de soortendiversiteit van waterstofproducerende bacteriën en methanogenen in C4, C6 en C7 hoger dan die in C2.
de omgevingsomstandigheden in het grondwater zullen rechtstreeks van invloed zijn op de groei en het metabolisme van micro-organismen. De pH-waarde van het grondwater in de kolenlaag is over het algemeen neutraal, maar de pH-waarde varieert tussen 6,5 en 8,4 in zandsteen gebroken aquifer in C4 gebied nr. 21 kolenlaag en het zoutgehalte is 1,0 g L−1. In de directe aquifer laag van de No. In het C6-gebied ligt de pH tussen pH 6,8 en 8,0 en het zoutgehalte is 0,7 g L−1. De pH-waarde van het grondwater in het C7-gebied is 6,1-7,3 en het zoutgehalte is 1,25 g L−1. De pH-waarde in C4, C6 en C7 is dicht bij neutraal en de graad van mineralisatie is laag, waar de micro-organismen gemeenschap betere groei, hogere overvloed en hogere diversiteit heeft. Daarnaast zijn het zoutgehalte van het grondwater en de ionensamenstelling nauw gerelateerd aan de anaerobe reductieomgeving van de kolenlaag. Bijvoorbeeld, SO42− wordt gebruikt om de gesloten omstandigheden van het grondwater te evalueren, en HCO3− is het product van de anaërobe ontzwaveling reactie van SO42−, dus hoge HCO3-kan worden gebruikt als een teken van een goede afdichting en sterke reductie van kolenbed grondwater . De waterchemie in het C4-gebied is HCO3 * SO4-Ca * Mg, de waterchemie in het C7-gebied is vergelijkbaar met het C4–gebied, HCO3·SO4-Ca·Na, en biedt een relatief gesloten anaerobe omgeving. In dit geval, is de chao1-index van waterstof-producerende bacteriën in C4 148, is de Shannon-index 1.52; de Chao1-index van methanogenen is 368, de Shannon-index is 2,35. De Chao1-index van waterstofproducerende bacteriën in C7 is 458, de Shannon-index is 1,98, de Chao1-index is 256 en de Shannon-index is 2,47. In C6 is de waterchemie SO4·HCO3–K·Na en is SO42− dominant, terwijl de Chao1-index van waterstofproducerende bacteriën in C6 472 is, de Shannon-index 1,56, de Chao1-index van methanogenen 384 en de Shannon-index 1,08. Gegevens tonen aan dat de diversiteit in C6 iets lager is dan C4 en C7.
sommige waterstofproducerende bacteriën en methanogenen werden gedetecteerd in C8-en C9-gebieden in het gebied waar geen biomethaan werd gevonden. Het was ook opmerkelijk dat de grondwateromstandigheden in deze twee gebieden vergelijkbaar zijn met die in de bovengenoemde biogene methaangebieden, die zich in de runoffzone van het grondwater bevinden en dat de grondwateraanvulling meer in staat is om organisch materiaal naar de kolenlaag te transporteren, zodat een groot aantal bacteriën groeit en zich vermenigvuldigt, wat een van de redenen is voor de grotere soortendichtheid en diversiteit van C8 en C9.
temperatuur
temperatuur en sporenmetaalelementen beïnvloeden de abundantie en diversiteit van microbiële gemeenschappen door zowel de groei als het metabolisme van micro-organismen en hun metabole omgeving direct te veranderen. Vanuit microbiologisch oogpunt is optimale temperatuur dus een van de belangrijkste factoren die de groei en het metabolisme van micro-organismen beïnvloedt. Figuur 5b laat zien dat de temperatuur een relatief zwakke invloed uitoefent op de overvloed en diversiteit van methanogenen, ook al bestaan waterstofproducerende bacteriën binnen een smalle ecologische amplitude en zijn gevoelig voor temperatuurveranderingen. Deze variabele is gecorreleerd met de dichtheid en diversiteit van de soorten, en de resultaten van deze studie tonen aan dat de temperatuur van de kolenlaag (d.w.z. tussen 25 en 27 °C) positief gecorreleerd is met de dichtheid van de bacteriële populatie. Bij C8 was de temperatuur 27,2 °C, de hoogste temperatuur die in deze studie werd geregistreerd. De Chao1-index van waterstof-producerende bacteriën was 510 en de Shannon-index was 2,61, ook de hoogste onder steekproeven (C1-C8). De laagste temperatuur, 24,9 °C, werd gevonden bij C1, waar de Chao1-index van bacteriële gemeenschap de laagste was. De abundantie en diversiteit van microbiële soorten neemt toe met de temperatuur in C3 > C6 > C2 > C7 > C5 > C4. Geothermische gradiëntafwijkingen bij C9 en C10 veroorzaakten veel hogere temperaturen; de omgevingstemperaturen bij C9 en C10 waren respectievelijk 34,60 °C en 40,10 °C. De Chao1-index van waterstof-producerende bacteriën in C9 was 176, en de Shannon-index was 1,28; de Chao1-index is 237, Shannon ‘ s index is 1,58. In vergelijking met de eerste acht gebieden is de overvloed en diversiteit iets afgenomen. Hier kunnen zowel waterstofproducerende bacteriën als methanogenen groeien en zich voortplanten bij de omgevingstemperatuur.
microbiële syntrofe interacties
in het extreme milieu van de kolenlaag worden consortia van bacteriën gevormd tussen de micro-organismen in de kolenlaag. Door de uitwisseling van metabolieten en micro-milieu gecontroleerde symbiose behouden concurrentie en middelen de specifieke functies van de microbiële gemeenschap, die de biomethaanproductieroute in de kolenlaag bepaalt. Methanothrix, dat azijnzuur omzet in methaan, is het dominante geslacht in de methaangemeenschap van het C1-gebied. De bacteriën geassocieerd met Alkalibaculum en Desulfosporosinus zijn homoacetogene bacteriën die H2 als elektronendonor gebruiken om azijnzuur te produceren. Zij zijn de belangrijkste concurrenten voor hydrogenotrofe methanogenen en bieden ook metabolisch substraat voor methanogenen. Waterstof-producerende bacteriën zoals Clostridium en Tissierella ook azijnzuur, en dus de hoge overvloed van waterstof-producerende bacteriën biedt een rijk metabolisch substraat voor Methanothrix. Samen zijn de methanogenen en de waterstof-producerende bacteriën in syntrofische interactie en de methaanproductie route in dit gebied wordt bepaald door de afbraak van azijnzuur. De methanogenen in C2, C4 en C6 zijn voornamelijk hydrogenotrofe methanogenen. Hydrolytische fermentatiebacteriën en acetogenen dragen beide bij aan de productie van azijnzuur en H2. Zij produceren ook enzymen, cofactoren en metabolische signalen om de waterstofproductie te regelen. Bovendien concurreren homoacetogene bacteriën en acetogenen niet op deze gebieden. Hydrogenotrofe methanogenen kunnen methaan produceren uit CO2 en H2 geproduceerd in de vorige fase. Daarom worden de metabolische wegen in deze gebieden hoofdzakelijk gebruikt voor H2, formate, en andere substanties.
meer dan 99% van het C3-gebied bevat methylotrofe methanogenen, zoals: Methanolobus. Brevibacter, Paenibacillus, Brochothrix en Lactococcus. Uit eerdere studies is gebleken dat methoxyaromatische verbindingen (een belangrijk deel van lignocellulose) worden afgebroken om methanol en andere stoffen te produceren . Micro-organismen in dit gebied kunnen de lignocellulose-achtige materie van steenkool degraderen om grondstoffen te leveren aan methylotrofe methanogenen. Deze eenvoudige microbiële gemeenschap kan niet voldoende substraten bieden voor methanogenen die H2 consumeren. De biomethaanproductie op dit gebied is gebaseerd op het verbruik van methylverbindingen.
Staphylococcus werd ook gedetecteerd in het C3-gebied. Onlangs werd Staphylococcus AntiMn-1 geïsoleerd uit diepzeesedimenten in het Clarion-Clippertongebied met een hoog mangaangehalte. Het bevatte genen met een hoge weerstand tegen mangaan, waarvan wordt gedacht dat het een aanpassing aan het mariene sedimentaire milieu . Het gehalte aan zware metalen in het C3-gebied is relatief hoog. Het kan zijn dat de omgeving van de kolenlaag de expressie van resistentiegenen effectief kan induceren, die antagonistische en ontgiftende effecten kunnen hebben op het transport en de toxiciteit van zware metalen binnen micro-organismen. Staphylococcus in dit gebied kan resistentiegenen bevatten om zich aan te passen aan de omgeving van de kolenlaag, zodat het ook kan deelnemen aan het fermentatiemetabolisme van steenkool. Er zijn veel verschillende soorten methanogenen in C7, en de waterstof-producerende bacteriën worden gedomineerd door Clostridium, Bacillus, Citrobacter, en andere anaëroben, die substraten voor acetoclastische methanogenen en ook H2, CO2, en formiaat voor hydrogenotrofe methanogenen. Verder vermindert ophoping van azijnzuur sulfaatreducerende bacteriën, waaronder Desulfosporosinus en Desulfitobacterium. SRB heeft een sterkere affiniteit voor azijnzuur dan acetoclastische methanogenen, maar ze concurreren niet met methylotrophic methanogenen voor bepaalde substraten, zoals methanol. Zo kunnen de metabolismen van zowel sulfaatreducerende bacteriën als methanogenen gelijktijdig op dit gebied doorgaan . Het metabolisme in C7 werd gedomineerd door methylotrofe methanogenen, gevolgd door azijnzuurfermentatie en vervolgens koolstofdioxide reductie.