DISCUSSION
we bepaalden hier de genoomgroottes van 25 chironomide soorten; deze waarden kwamen overeen met de genoomgroottes van de drie soorten die eerder in de literatuur werden gerapporteerd (Petitpierre, 1996; Schmidt-Ott et al., 2009), die tussen 0.13 en 0.21 PG DNA werden samengesteld. De resultaten van deze studie toonden aan dat de genoomgroottes van chironomiden uniform zeer klein waren (Fig. 3, Tabel 1). De verkregen gemiddelde C-waarde voor de Chironomidae familie is 0,13 + 0.03 PG DNA, dat duidelijk kleiner is dan D. melanogaster genoomgrootte. De kleinste genoomgrootte onder insecten is geregistreerd in de Strepsiptera Caenocholax fenyesi met ongeveer 0,11 pg DNA (overeenkomend met ongeveer 108 Mb), hoewel de Hessische vlieg Mayetiola destructor ook werd gesuggereerd om een nog kleinere genoomgrootte van 0,09 pg te hebben (Johnston et al., 2004; Gregory, 2014). Hier rapporteren we ten minste drie soorten met kleinere genomen dan elk bekend insect: C. tsushimensis, Diamesa japonica en Hydrobaenus tsukubalatus, met C-waarden van respectievelijk 0,07 pg, 0,08 PG en 0,08 pg (Tabel 1). Voor zover wij weten, C. tsushimensis (Fig. 1B) Kan nu worden beschouwd als het hebben van de kleinste genoomgrootte van om het even welk bekend insect, met een c-waarde van 0.07 PG DNA, die ruwweg aan 68 Mb overeenkomt. Deze waarde is congruent met de genoomgrootte van een verwante soort, Clunio marinus, die eenmaal werd geschat op 95 Mb en recenter op 87,2 Mb (Kaiser and Heckel, 2012; Tobias S. Kaiser, personal communication).
onlangs werd het genoom van de Antarctische midge, B. antarctica, gesequenced (Kelley et al., 2014). De auteurs beweerden dat de genoomgrootte van deze soort, geschat op 89.5-105 Mb, was de kleinste onder alle insecten. We laten hier echter zien dat deze genoomgrootte binnen het normale bereik van chironomiden-waarden ligt. De auteurs suggereerden ook dat de kleine genoomgrootte van B. antarctica een aanpassing aan extreme koude omgeving was. Onze gegevens kunnen deze hypothese bevestigen, aangezien D. japonica, een alpensoort uit de onderfamilie Diamesinae ook een kleine genoomgrootte heeft. Een andere verwante Diamesinae, de Himalaya midge Diamesa sp. bleek actief te zijn bij temperaturen zo laag als -16°C (Kohshima, 1984). Telmatogeton japonicus, die zich ook bij zeer lage temperaturen kan ontwikkelen (Danks, 1971; Sunose en Fujisawa, 1982), vertoont echter een relatief grote genoomgrootte, vergeleken met het gemiddelde van de chironomiden. Hierbij moet rekening worden gehouden met een fylogenetische invloed op de grootte van het genoom, aangezien B. antarctica behoort tot de orthocladiinae subfamilie, die gemiddeld kleine genoomgroottes vertoont. De soorten C. tsushimensis en Hydrobaenus tsukubalatus, met de kleinste genoomgrootten onder de chironomiden, behoren ook tot de orthocladiinae onderfamilie. Aldus, de kleine genoomgrootte van B. antarctica kan gewoon een voorouderlijke eigenschap zijn, niet het resultaat van extreme koude aanpassing.
P. Vanderplanki is de enige insectensoort waarvan bekend is dat hij anhydrobiose kan bereiken in een volledig gedehydrateerde toestand (Fig. 1D) en herstellen na rehydratatie (Hinton, 1951, 1960a; Watanabe, 2006; Cornette en Kikawada, 2011). Onze onderzoeksgroep publiceerde onlangs een vergelijkende analyse van de genoomsequenties van het desiccation-tolerante Polypedilum vanderplanki en de desiccation-sensitive congeneric Polypedilum nubifer (Gusev et al., 2014). Uit de vergelijking bleek dat beide soorten vergelijkbare genoomgrootten hadden en dat slechts een beperkte set van gedupliceerde genclusters gerelateerd was aan anhydrobiose in P. Vanderplanki (Gusev et al., 2014). De genoomgroottes die uit de genoomassemblage werden afgeleid, waren 104 Mb voor P. Vanderplanki en 107 Mb voor P. nubifer en deze waarden verschilden niet aanzienlijk van de genoomgroottes die in deze studie voor deze soorten werden gemeten, die overeenkwamen met ongeveer 98 Mb. P. vanderplanki en P. het genoom van nubifer vertoonde een vergelijkbaar laag percentage DNA-herhalingen en er werd slechts een klein aantal transposeerbare elementen gevonden, vergeleken met andere dipteran-soorten. Deze vermindering van het aandeel van transposeerbare elementen werd ook waargenomen in de Antarctische midge, B. antarctica (Kelley et al., 2014). De gemiddelde intron lengte was ook aanzienlijk verminderd in de Antarctische midge en in zowel P. Vanderplanki als P. nubifer (Gusev et al., 2014). Sinds de uitdroging gevoelige en bevriezen intolerante P. nubifer deelt deze kenmerken met de Antarctische midge en de anhydrobiotische midge, een laag aandeel van transposeerbare elementen en een korte intron-lengte kunnen een kenmerk zijn dat door alle chironomiden wordt gedeeld en kunnen niet rechtstreeks verband houden met de tolerantie voor extreme omgevingen, zelfs als een adaptief effect niet kon worden uitgesloten.
Fig. 3.
fylogenie van Diptera met geschatte C-waarden. De Chironomide familie is in het zwart afgebeeld. De streep toont de genoomgrootte van D. melanogaster. De fylogenetische cladogram is een consensus van verschillende Chironomide en Dipteran bomen (Yeates and Wiegmann, 1999, 2005; Saether, 2000; Cranston et al., 2012). Fouten balken tonen standaardafwijking. De grootte van het genoom zonder fout bars werd verkregen uit vorige studies die in de Database van de dierlijke Grootte van het genoom worden vermeld (Gregory, 2014).
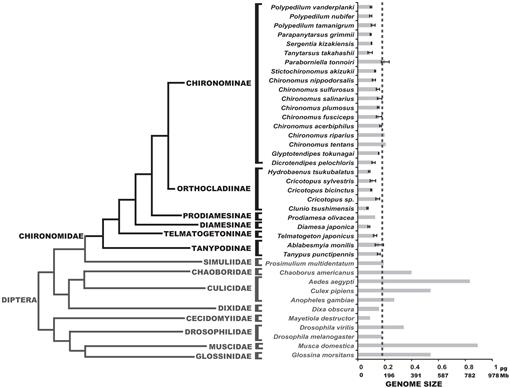
Fig. 4.
relatie tussen lichaamsgrootte en genoomgrootte van de chironomide-soorten die in dit onderzoek zijn onderzocht. A) voor elke soort wordt de gemiddelde totale lichaamslengte aangegeven in millimeters en de gemiddelde C-waarde uitgedrukt in pg DNA. Slechts een zwakke positieve correlatie (r2 = 0,171, n = 25), gematerialiseerd door een vaste lijn, werd waargenomen tussen de twee variabelen. (B) op geslachtsniveau werden middelen van dezelfde waarden getransformeerd door de methode van fylogenetisch onafhankelijke contrasten, gebaseerd op de fylogenie van Cranston et al. (2012). De correlatie tussen deze getransformeerde contrasten, gematerialiseerd door een vaste lijn, was niet significant (r2 = 0,118, n = 13). Gegevensanalyse werd uitgevoerd met Prism 4.0 software voor Macintosh (GraphPad Software, Inc.).

wat kunnen de adaptieve eigenschappen zijn die mogelijk geassocieerd worden met de kleine genoomgroottes van de Chironomidae familie? In feite, zijn verscheidene ontwikkelings en ecologische factoren voorgesteld om met genoomgrootte te correleren. Ten eerste is in veel taxa een positieve correlatie gevonden tussen de grootte van het genoom en de lichaamsgrootte en deze correlatie verschijnt als een relatief algemeen verschijnsel (Gregory, 2005). Deze correlatie is ook gemeld bij muggen (Ferrari en Rai, 1989), maar is niet altijd duidelijk bij andere insectentaxa (Gregory en Hebert, 2003; Ardila-Garcia en Gregory, 2009). Binnen de chironomiden vonden we slechts een zwakke positieve correlatie tussen genoomgrootte en lichaamsgrootte (Fig. 4A). De gemiddelde C-waarde voor het geslacht Chironomus (0,15 pg) was hoger dan die voor het geslacht Polypedilum (0,10 pg), en dit verschil kan worden toegeschreven aan de over het algemeen grotere lichaamsgrootte van Chironomus-soorten. Nochtans, kon de phylogenetic afstand tussen beide geslachten dit verschil van genoomgrootte ook verklaren. Daarom corrigeerden we onze gegevens op genus niveau met fylogenetisch onafhankelijke contrasten analyse (Garland and Adolph, 1994; Garland et al ., 2005) en als gevolg daarvan bleek de correlatie tussen genoomgrootte en lichaamsgrootte niet significant te zijn (Fig. 4B). Het nauwe bereik van de Genoomgrootten van Chironomidae (slechts twee—drievoudig) en de gemiddelde kleine lichaamsgrootte van deze soorten (de meeste niet meer dan een paar millimeter) maken het waarschijnlijk moeilijk om een correlatie te vinden tussen beide eigenschappen binnen de Chironomidae familie.
bij insecten werd een andere correlatie gesuggereerd tussen de grootte van het genoom en de complexiteit van de ontwikkeling. Terwijl hemimetabole insecten, met alleen geleidelijke nimfale vervelling, een breed scala aan genoomgroottes vertonen (C-waarden van 0,18 tot 16.93 pg), holometabole insecten met hun complexe metamorfose hebben C-waarden beperkt binnen een vermeende 2 pg drempel, voor de meeste van hen (Gregory, 2002, 2005). De invloed van ontwikkelingscomplexiteit op genoomgrootte is hier duidelijk omdat holometabolous insecten intensieve morfologische remodellering tijdens de beperkte tijd van metamorfose ondergaan en een kleine genoomgrootte voordelen voor het bereiken van celdelingen aan een hoog tarief voorstelt. Chironomidae zijn holometabole insecten en hun metamorfose is extreem snel. Hun pupale stadium kan slechts één dag duren, of zelfs enkele uren voor bepaalde soorten (Cranston, 1995c). C. tsushimensis vertoont bijvoorbeeld een complexe metamorfose met uitgesproken seksueel dimorfisme (Fig. 1 ter).
de relatie tussen de grootte van het genoom en de snelheid van de celdeling beïnvloedt ook de ontwikkeling. Bijvoorbeeld, werd de snelle levenscyclus van bladluizen voorgesteld om aan hun kleine genoomgrootte (Ma et al., 1992). Aldus, zouden de kleine genoomgrootte een voordeel voor chironomid species moeten vormen die zich snel in tijdelijke milieu ‘ s ontwikkelen. Chironomiden van het geslacht Clunio kunnen zich ontwikkelen tot tijdelijke getijdenpoelen en tonen ook gesynchroniseerde opkomst van de volwassenen in relatie tot de maancyclus en tijdens het korte venster van het laagwater (Kaiser and Heckel, 2012). Om dit te bereiken is een hoge ontwikkelingsgraad nodig. Merk op dat kleine polychaete anneliden die zich snel ontwikkelen in vergelijkbare interstitiële omgevingen kleinere genoomgrootten vertonen dan macrobenthische soorten (Gambi et al., 1997). Voor Antarctische muggen, zoals B. antarctica of eretmoptera murphyi (Lee et al., 2006; Worland, 2010), larvale ontwikkeling duurt twee jaar, meestal gearresteerd in een bevroren toestand, maar metamorfose en voortplanting moet plaatsvinden tijdens de zeer korte zomerperiode. In dit geval, is een hoog ontwikkelingstarief nodig en dit kan door het kleine genoom van B. antarctica worden vergemakkelijkt. Dit fenomeen werd geïllustreerd door een studie over angiosperm planten, waaruit blijkt dat de soorten in staat om hun hele levenscyclus te voltooien tijdens de korte Antarctische zomer presenteerde de kleinste genoomgroottes (Bennett et al., 1982). Een andere extreme, de anhydrobiotische midge P. Vanderplanki is aangepast aan kortstondige rotspoelen. Ook hier kan een hoge ontwikkelingsgraad in een tijdelijke habitat in verband worden gebracht met de kleine genoomgrootte van deze soort. Echter, de Australische soort P. tonnoiri, die ook wordt blootgesteld aan uitdroging op soortgelijke rotsbaden (Jones, 1975; Adams, 1985; Frouz et al., 2003), toont een relatief grote genoomgrootte voor een chironomide (Fig. 3 en Tabel 1).
abiotische spanningen beïnvloeden waarschijnlijk ook de integriteit van het DNA en als gevolg daarvan de grootte van het genoom. Bijvoorbeeld, larven van P. vanderpianki ervaren enorme DNA-schade na anhydrobiose en efficiënte DNA-reparatie vindt plaats gedurende de paar dagen na rehydratie (Gusev et al., 2010). In planten werd een negatieve correlatie gevonden tussen de grootte van het genoom en de tolerantie voor straling (Bennett and Leitch, 2005) en het mutatiepercentage was ook lager bij soorten met een kleinere genoomgrootte (Abrahamson et al., 1973). Bijgevolg kan de kleine genoomgrootte van P. Vanderpianki een voordeel zijn om de accumulatie van schadelijke mutaties te voorkomen tijdens de DNA-herstelgebeurtenissen geassocieerd met cycli van uitdroging en rehydratie. Als mariene soort wordt C. tsushimensis ook blootgesteld aan een hoge zoutstress, wat dodelijk is voor de meeste insectensoorten. Het hoge zoutgehalte en andere abiotische spanningen zijn gekend om intracellular reactieve zuurstofspecies (ROS) te produceren, die als belangrijkste bron van DNA-schade werden voorgesteld (Franca et al., 2007; Gill and Tuteja, 2010). Dit kan ook worden gerelateerd aan kleine genoomgrootte. Echter, andere mariene en hoog zoutgehalte tolerante soorten, zoals T. japonicus of Chironomus salinarius hebben niet genoom maten zo klein als C. tsushimensis. Zuur-tolerante soorten, zoals Chironomus sulfurosus, Chironomus acerbiphilus of Polypedilum tamanigrum (Fig. 1C) kan overleven in warmwaterbronnen met een pH van 1,4 (Doi et al., 2004; Takagi et al., 2005). Ook hier kon geen duidelijke correlatie tussen tolerantie voor zure stress en kleine genoomgrootte worden waargenomen en de fylogenetische invloed bleek belangrijker, met kleinere genoomgrootte voor het geslacht Polypedilum en grotere genoomgroottes in het geslacht Chironomus.De kleine genoomgrootten die bij chironomiden worden waargenomen, zijn waarschijnlijk het resultaat van een voorouderlijke vermindering van het aantal transposeerbare elementen en van de lengte van introns in hun genoomstructuur. Bij de Chironomidae-familie verschilden de genoomgroottes naar gelang van de fylogenie op subfamilie – niveau en op genus-niveau. Zoals afgeleid uit correlaties in verschillende taxa, zou de kleine genoomgrootte van chironomids theoretisch een preadaptatie aan onstabiele en extreme milieu ‘ s door hoge ontwikkelingstarief en lage mutatietarief kunnen hebben gevormd. We hopen dat deze studie toekomstige genoomprojecten van chironomiden zal aanmoedigen en dat vergelijkende genomica binnen dit taxon zal helpen om te begrijpen hoe de genomen van chironomiden interageren met hun omgeving.