analiza czynników wpływających na strukturę społeczności mikrobiologicznej
w celu dalszego zbadania możliwych zależności między czynnikami środowiskowymi a wariancją społeczności opracowano analizę RDA (rys. 7). W sumie osiem czynników środowiskowych, w tym pierwiastki śladowe Fe, Co, Ni, temperatura, zasolenie, głębokość, wilgotność i RO. W miejscu pobierania próbek zmierzono głębokość i temperaturę zbiornika, a inne informacje uzyskano z danych geologicznych lokalnych kopalń (zob. tabela 2). Dane pokazane na Rys. 7A ujawniło, że kompozycje społeczności bakteryjnej Znalezione w tym badaniu miały znaczący wpływ na Fe, Ni, wilgoć, zasolenie i RO. Wszystkie społeczności, inne niż C4, C7, C10, są pozytywnie skorelowane z RO; C4, C7 I C10 pozytywnie skorelowane z Fe, Ni i wilgocią. Co jest wymagane dla koenzymu metylo-transferazy m, który jest ważnym enzymem w biochemicznym metabolizmie metanogenów ; w związku z tym wpływ Co na środowisko archaealne jest większy niż w przypadku społeczności bakteryjnej. Pierwiastki Fe, Co i Ni, a także wilgoć, okazały się najważniejszymi czynnikami środowiskowymi, a następnie RO i zasolenie w społeczności archaealnej. Istnieje znacząca dodatnia korelacja między co a Wspólnotami C1, C8, C9, C10. Wszystkie społeczności z wyjątkiem C2, C4, C6 I C5 były ujemnie skorelowane z zasoleniem (Fig. 7b).

RDA (analiza redundancji) oparta na poziomie bakterii (a) i archeonów (b)z czynnikami środowiskowymi złoża węgla i charakterystyką węgla. Długość impact factor jest dłuższa, udział impact jest wyższy, a odwrotnie, gdy impact factor jest krótszy, udział impact jest lżejszy. Gdy czynnik środowiskowy jest Ostro nachylony z próbką, istnieje korelacja dodatnia, a gdy czynnik środowiskowy i kąt próbki są rozwarte, istnieje korelacja ujemna
stopień węgla
„skok” węgla odnosi się do serii zmian fizycznych i chemicznych pod wpływem temperatury i ciśnienia węgla w historii geologicznej. W ten sposób węgiel przeszedł proces stopniowej do nagłej zmiany. Cztery skoki odpowiadają RO 0,6, 1.3, 2,5 i 3,0%. Niezależnie od archaealnej czy bakteryjnej społeczności, ranga węgla ma pewien wpływ na różnorodność i obfitość bakterii. Wraz ze wzrostem rangi węgla, zarówno w społecznościach archaealnych, jak i bakteryjnych, różnorodność społeczności wykazuje pewną ogólną tendencję spadkową(rys. 8). Ponadto mikroorganizmy mogą wpływać na skład węgla kontrolowanego przez szeregi węglowe. Węgle średniej i niskiej rangi zawierają duże ilości substancji wyewoluowanych przez rośliny w Grupie 1, która zawiera wiele substancji wyewoluowanych przez rośliny. Tutaj jest wyższa zawartość wodoru, tlenu i azotu; a składniki odżywcze wymagane przez bakterie są obfite. W procesie koalifikacji substancje organiczne wytwarzają dużo wilgoci i ciekłych węglowodorów. Jednocześnie obfite są również łańcuchy boczne wodoru i tlenu zawarte w węglu. Te płynne i stałe substancje stanowią podstawę życia dla bakterii. W rezultacie bogactwo i różnorodność bakterii produkujących wodór i metanogeny w węglu w tym regionie są stosunkowo wysokie. Wraz ze wzrostem Ro, zawartość łańcucha bocznego wodoru i tlenu w węglu jest drastycznie zmniejszona, a składniki dostępne dla mikroorganizmów są również zmniejszone. W związku z tym liczebność i różnorodność gatunkowa zbiorowisk bakteryjnych i archaealnych w grupie 2 i grupie 3 jest ogólnie zmniejszona. Do tej pory stwierdzono, że w przyrodzie poziomy biogennego metanu z pokładów węgla mają współczynnik odbicia 2,0% (C4 Hebi). Po RO > 2,5% związki organiczne, które można przekształcić w małe cząsteczki, były bardzo rzadkie, ale w grupie 4 odnotowano większą różnorodność i obfitość. Spekulujemy, że składniki odżywcze wprowadzane przez wody gruntowe w tym czasie są dostępne dla rozmnażania bakterii. Składniki odżywcze wytworzone przez wody gruntowe w różnych regionach i w różnych porach roku mogły przyczynić się do różnorodności gatunków. Jednym z powodów większej różnorodności flory w grupie 4 może być to, że obszar C8 Jiaozuo Jiulishan ma lepsze warunki spływu wód gruntowych i silniejsze Ładowanie. Może transportować składniki odżywcze dla flory, więc różnorodność i obfitość są wyższe niż Grupa 2. Warto zauważyć, że różnorodność i obfitość zbiorowisk archaealnych jest w pewnym stopniu negatywnie skorelowana z szeregami węglowymi. Jednak liczebność gatunków w zbiorowiskach bakteryjnych jest pozytywnie skorelowana z szeregiem węglanów, a różnorodność wykazuje tendencję spadkową. Wraz ze wzrostem Rang węglowych niektóre grupy bakterii stopniowo dostosowują się do środowiska różnych rang węglowych i mogą rosnąć i rozmnażać się w dużych ilościach, a metanogeny są trudne do przystosowania do rang węglowych.
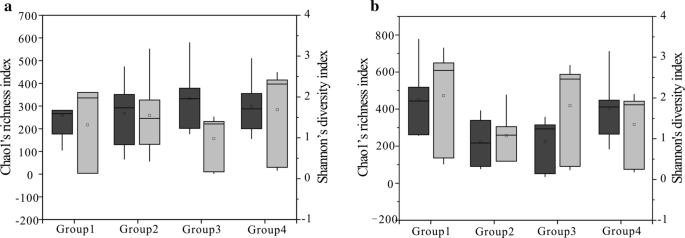
indeks Chao1 (ciemnoszary) i Shannona (jasnoszary) dla czterech grup (próbki węgla podzielono na cztery grupy według wartości RO dla wspólnoty bakteryjnej (a) i wspólnoty archaicznej (b), Grupa 1 oznacza wartość mniejszą niż 0,6%, Grupa 2 oznacza wartość od 0,8 do 1,1%, Grupa 3 oznacza wartość od 1,4 do 1,8%, Grupa 4 oznacza wartość od 2,67 do 3,15%) pochodzących z regionów. 25. I 75. percentyle są wskazywane przez zewnętrzne krawędzie pudełek, podczas gdy wartości maksymalne i minimalne są pokazywane przez końce wąsów, a mediana przez poziomą linię w każdym pudełku
pierwiastki śladowe
pierwiastki śladowe mogą promować wzrost mikroorganizmów w pewnym zakresie, w którym komórka utrzymuje homeostazę pierwiastków poprzez regulację metaboliczną. Pierwiastki śladowe mogą również występować w różnych enzymach, które mogą być wchłaniane i wykorzystywane przez mikroorganizmy w procesie metabolizmu beztlenowego, co ma wpływ na strukturę wspólnotową bakterii produkujących wodór i metanogenów (Tabela 3).
Fe i Ni mają większy wpływ na bakterie produkujące Wodór niż Co . Fe i Ni mogą uczestniczyć w syntezie i metabolizmie hydrogenaz i innych metaloenzymów w mikroorganizmach. W miarę jak zawartość Fe i Ni wzrasta w pewnym zakresie, podobnie jak obfitość i różnorodność populacji bakterii produkujących Wodór. Zawartość Fe i Ni w C7 jest znacznie wyższa niż w innych regionach, a praca ta wykazała, że Clostridium są bakteriami produkującymi Wodór. To odkrycie wskazuje, że nadmierne poziomy pierwiastków śladowych mogą mieć toksyczny wpływ na wzrost mikroorganizmów i hamować aktywność metaloenzymów. Poziomy Fe w C4, C6 i C9 nie różniły się znacząco i były stabilne przy 3500 mg Kg-1 (Fig. 9). Względna obfitość Ni w trzech obszarach wynosi C6 > C9 > C4, co odpowiada porządkowi obfitości (również C6 > C9 > C4), ale porządek różnorodności to C6 > C4 > C9. W zbiorowiskach C4, C6 i C9 wykryto przedstawicieli rodzajów: Clostridium, Klebsiella, Enterobacter i Citrobacter, w tym tych o większej obfitości i zróżnicowaniu niż w innych regionach.

zawartość pierwiastków śladowych w Fe, Co, Ni w próbkach węgla
w społeczności archaealnej wpływ Fe, Co i Ni na metanogeny jest jeszcze ważniejszy. Co jest kluczowym elementem w syntezie metanogennego koenzymu F430, a zawartość Co w pierwszej trójce wynosi C8 > C7 > C10, przy czym gatunki i liczebność metanogenu to C8 > C7 > C10. Zawartość Co jest w pewnym stopniu pozytywnie skorelowana z obfitością i różnorodnością metanogenów. Chociaż zawartość Fe w C7 jest znacznie wyższa niż w innych regionach, nie wpływa na dystrybucję metanogenów w regionie. Istnieje tylko kilka rodzajów metanogenów, które mogą zawierać Fe – we wcześniejszych badaniach odkryto tylko jeden gatunek, nazwany Methanothermobacter. Obecność monoferrycznej hydrogenazy w metanogenach M. marburgensis katalizuje odwracalną reakcję metenylo-H4MPT+ i H2 w celu wytworzenia metyleno-H4MPT i h+, wytwarzając metan z CO2 i H2 . Podobne enzymy mogą również zawierać metanogeny wykorzystujące metabolizm hydrogenotropowy. Ponadto spekuluje się, że duża część metanogenów metylotroficznych zawiera takie enzymy, z wyjątkiem Metanokulli i Metanobakterii. Spekuluje się, że w metanogenach metylotroficznych może występować metaloenzym związany z Fe.
warunki gruntowe
woda gruntowa bezpośrednio lub pośrednio stanowi ekologiczną podstawę wzrostu i metabolizmu ekstremofilów w pokładie węgla. Z jednej strony, groundwater recharge dostarcza dużych ilości składników odżywczych dla społeczności bakteryjnych i archaealnych; z drugiej strony, warunki środowiskowe wód gruntowych (Eh, pH, zasolenie, skład jonów i pierwiastki śladowe) bezpośrednio wpływają na wzrost drobnoustrojów i aktywność enzymów metabolicznych. Warunki środowiskowe wód podziemnych są bezpośrednio związane z wykorzystaniem i degradacją węgla, a mikroorganizmy znajdujące się w pokładie węgla wykazują różne struktury społeczności i cechy funkcjonalne.
drobnoustrojowe substraty odżywcze są zazwyczaj rozpuszczane. Strefa odpływu w obszarze górniczym może umożliwiać przetrwanie pokładów węgla. Zbiorniki o wysokiej przepuszczalności mają pozytywny wpływ na wzrost i reprodukcję bakterii produkujących wodór i metanogeny, natomiast Metamorfizm ma znaczący negatywny wpływ na przepuszczalność węgla w zbiornikach węglowych . Na obszarach o biogennym CBM społeczności C2, C4, C6 I C7 zostały dobrze udokumentowane. Społeczności te należą do niskiej i średniej rangi węgla, porowatość węgla jest stosunkowo wyższa niż węgiel wysokiej rangi, wody gruntowe mogą dostarczać składniki odżywcze drobnoustrojom w pokładie węgla w czasie. Obecna strefa rozwoju CBM w dorzeczu rzeki Powder w USA koncentruje się głównie w strefie spływu wód gruntowych. Stabilne gazowo dane izotopowe z płytkiej studni CBM w obszarze wydobywczym C6 potwierdziły również obecność biogenetycznego CBM w tym obszarze. Jednak dane dotyczące izotopów stabilnych gazowo z innej głębokiej studni CBM wskazywały, że CBM jest głównie termogeniczny. Wyniki te pokazują, że wraz ze wzrostem głębokości pochówku, warunki odpływu osłabną i trudno będzie transportować składniki odżywcze dla mikroorganizmu, co spowoduje zmniejszenie obfitości i różnorodności społeczności. Dach i Podłoga Nie 2 szew węglowy w obszarze C2 ma stosunkowo stabilne warstwy kamienia błotnego i gliny, co utrudnia bakteriom produkującym wodór i metanogeny w szwie węglowym uzyskanie płynnych składników odżywczych, a także ogranicza ich wzrost i metabolizm, a tym samym ich różnorodność i obfitość społeczności. Zauważ, że w tym obszarze wskaźnik Chao1 wynosi 240, a wskaźnik Shannona wynosi 1,38 W społeczności bakterii. Wskaźnik Chao1 metanogenu wynosi 82, wskaźnik Shannona 0,56. Piaskowiec szczelinowy dach wodonośny w rejonie C4 o nr 21 pokład węglowy ma lepsze warunki ładowania i wypełnia pokład węglowy wodą. Możliwe jest, że społeczność mikrobiologiczna doświadcza kumulacyjnych skutków wystarczającej dostępności różnych składników odżywczych, co wpływa na transport, w porównaniu z obfitością i różnorodnością społeczności mikroorganizmów w obszarze C2, która znacznie się poprawiła. W tym obszarze wskaźnik Chao1 bakterii produkujących Wodór wynosi 148, wskaźnik Shannona 1,52, wskaźnik Chao1 metanogenów 368, a wskaźnik Shannona 2,35. Warstwa wodonośna wypełniona piaskowcem o powierzchni C6 jest bezpośrednio wypełnioną wodą warstwą wodonośną złoża węgla nr 3. Rozwój pęknięć w warstwie i umiarkowana aqueosity również odgrywa aktywną rolę w obfitości i różnorodności społeczności. Tutaj wskaźnik Chao1 bakterii produkujących Wodór wynosi 472, wskaźnik Shannona 1,56, wskaźnik Chao1 metanogenów 384, a wskaźnik Shannona 1,08. Tak jest również w obszarze C7, pokład węgla nr 5 mA bezpośrednio pękniętą warstwę wodonośną o dobrych warunkach ładowania, wskaźnik obfitości bakterii produkujących Wodór wynosi 458, wskaźnik Shannona wynosi 1,98; wskaźnik Chao1 metanogenu wynosi 256, a wskaźnik Shannona 2,47. Dlatego różnorodność gatunkowa bakterii produkujących wodór i metanogenów w C4, C6 i C7 jest wyższa niż w C2.
warunki środowiskowe wód gruntowych bezpośrednio wpływają na wzrost i metabolizm mikroorganizmów. Wartość pH wód podziemnych z pokładu węgla jest na ogół neutralna, ale wartość pH waha się między 6,5 A 8,4 w piaskowcowych pękniętych warstwach wodonośnych w obszarze C4 nr 21, a zasolenie wynosi 1,0 g L−1. W bezpośredniej warstwie wodonośnej Nie 3 pokład węgla w obszarze C6, pH waha się od pH 6,8 do 8,0, a zasolenie wynosi 0,7 g L-1. Wartość pH wód podziemnych obszaru C7 wynosi 6,1-7,3, a zasolenie 1,25 G L-1. Wartość pH w C4, C6 i C7 jest zbliżona do neutralnej, a stopień mineralizacji jest niski, gdzie społeczność mikroorganizmów ma lepszy wzrost, większą obfitość i większą różnorodność. Ponadto zasolenie wód gruntowych i skład jonów są ściśle związane ze środowiskiem redukcji beztlenowej pokładu węgla. Na przykład SO42-służy do oceny zamkniętych warunków wód gruntowych, a HCO3− jest produktem beztlenowej reakcji odsiarczania SO42−, tak wysoka HCO3− może być stosowana jako znak dobrego uszczelnienia i silnej redukcji wód podziemnych z pokładu węgla . Chemia wody w obszarze C4 jest HCO3 * SO4-Ca * Mg, Chemia wody w obszarze C7 jest podobna do obszaru C4, HCO3 * SO4-Ca * Na i zapewnia stosunkowo zamknięte środowisko beztlenowe. W tym przypadku wskaźnik Chao1 bakterii produkujących wodór w C4 wynosi 148, wskaźnik Shannona wynosi 1,52; wskaźnik Chao1 metanogenu wynosi 368, wskaźnik Shannona 2,35. Wskaźnik Chao1 bakterii produkujących wodór w C7 wynosi 458, wskaźnik Shannona 1,98, wskaźnik Chao1 256, a wskaźnik Shannona 2,47. W C6 Chemia wody wynosi SO4 * HCO3-K * Na, a SO42-dominuje, podczas gdy wskaźnik Chao1 bakterii produkujących wodór w C6 wynosi 472, wskaźnik Shannona wynosi 1,56, wskaźnik Chao1 metanogenów wynosi 384, a wskaźnik Shannona wynosi 1,08. Dane pokazują, że różnorodność w C6 jest nieco niższa niż C4 I C7.
niektóre bakterie produkujące wodór i metanogeny zostały wykryte w obszarach C8 i C9 w obszarze, w którym nie znaleziono biometanu. Warto również zauważyć, że warunki gruntowe tych dwóch obszarów są podobne do tych w wyżej wymienionych obszarach biogennych metanu, które znajdują się w strefie spływu wód gruntowych, a Ładowanie wód gruntowych jest bardziej zdolne do transportu materii organicznej do pokładu węgla, dzięki czemu rośnie i namnaża się duża liczba bakterii, co jest jedną z przyczyn większej liczebności i różnorodności gatunków C8 i C9.
Temperatura
temperatura i pierwiastki śladowe wpływają na obfitość i różnorodność społeczności mikrobiologicznych, bezpośrednio zmieniając zarówno wzrost, jak i metabolizm mikroorganizmów oraz ich środowisko metaboliczne. Dlatego z mikrobiologicznego punktu widzenia optymalna temperatura jest jednym z najważniejszych czynników wpływających na wzrost i metabolizm mikroorganizmów. 5B pokazuje, że temperatura wywiera stosunkowo słaby wpływ na obfitość i różnorodność metanogenów, mimo że bakterie produkujące Wodór istnieją w wąskiej amplitudzie ekologicznej i są wrażliwe na zmiany temperatury. Zmienna ta jest skorelowana z liczebnością i różnorodnością gatunków, a wyniki badań pokazują, że temperatura pokładu węgla (tj. między 25 a 27 °C) jest dodatnio skorelowana z liczebnością populacji bakterii. W C8 temperatura wynosiła 27,2 °C, najwyższa temperatura odnotowana w tym badaniu. Wskaźnik Chao1 bakterii produkujących Wodór wynosił 510, A wskaźnik Shannona 2,61, również najwyższy wśród próbek (C1-C8). Najniższą temperaturę, 24,9 °c, stwierdzono w C1, gdzie indeks Chao1 społeczności bakteryjnej był najniższy. Obfitość i różnorodność gatunków drobnoustrojów wzrasta wraz z temperaturą w C3 > C6 > C2 > C7 > C5 > C4. Anomalie gradientu geotermalnego w C9 I C10 powodowały znacznie wyższe temperatury; temperatury otoczenia w C9 I C10 wynosiły odpowiednio 34,60 °C i 40,10 °C. Wskaźnik Chao1 bakterii produkujących wodór w C9 wynosił 176, a wskaźnik Shannona 1,28; wskaźnik Chao1 237, wskaźnik Shannona 1,58. W porównaniu do pierwszych ośmiu obszarów liczebność i różnorodność nieznacznie się zmniejszyły. Tutaj zarówno bakterie produkujące Wodór, jak i metanogeny mogą rosnąć i rozmnażać się w temperaturze otoczenia.
mikrobiologiczne interakcje syntroficzne
w ekstremalnym środowisku pokładu węgla, wśród mikroorganizmów pokładu węgla tworzą się konsorcja bakterii. Poprzez wymianę metabolitów i symbiozę kontrolowaną przez mikro-środowisko, konkurencja i alokacja zasobów utrzymują specyficzne funkcje społeczności mikrobiologicznej, która określa ścieżkę produkcji biometanu w pokładie węgla. Methanothrix, który przekształca kwas octowy w metan, jest dominującym rodzajem we Wspólnocie metanogenu w obszarze C1. Bakterie związane z Alkalibaculum i Desulfosporosinus są bakteriami homoacetogennymi, które wykorzystują H2 jako donor elektronów do produkcji kwasu octowego. Są głównymi konkurentami dla metanogenów hydrogenotropowych, a także dostarczają substratu metabolicznego dla metanogenów. Bakterie produkujące Wodór, takie jak Clostridium i Tissierella, również dostarczają kwasu octowego, a zatem wysoka obfitość bakterii produkujących Wodór zapewnia bogate podłoże metaboliczne dla Methanothrix. Razem metanogeny i bakterie produkujące Wodór znajdują się w interakcji syntroficznej, a szlak wytwarzania metanu w tym obszarze jest określany przez rozkład kwasu octowego. Metanogeny w C2, C4 i C6 to głównie metanogeny hydrogenotropowe. Bakterie fermentacji hydrolitycznej i acetogeny przyczyniają się do produkcji kwasu octowego i H2. Wytwarzają również enzymy, kofaktory i sygnały metaboliczne, aby regulować produkcję wodoru. Ponadto bakterie homoacetogenne i acetogeny nie konkurują w tych obszarach. Metanogeny hydrogenotropowe mogą wytwarzać metan z CO2 i H2 wytworzonych w poprzednim etapie. Dlatego szlaki metaboliczne w tych obszarach są wykorzystywane głównie dla H2, mrówczanów i innych substancji.
ponad 99% obszaru C3 zawiera metanogeny metylotroficzne, takie jak: Metanolobus. Brevibacter, Paenibacillus, Brochothrix i Lactococcus. Wcześniejsze badania wykazały, że związki metoksyaromatyczne (ważna część lignocelulozy) są rozkładane do produkcji metanolu i innych substancji . Mikroorganizmy w tym regionie mogą degradować lignocelulozową materię węgla, aby zapewnić zasoby metanogenom metylotroficznym. Ta prosta społeczność mikrobiologiczna nie może zapewnić wystarczającej ilości substratów dla metanogenów, które zużywają H2. Ścieżka produkcji biometanu w tym obszarze opiera się na zużyciu związków metylowych.
gronkowce Wykryto również w obszarze C3. Ostatnio Staphylococcus AntiMn-1 wyizolowano z osadów głębinowych w rejonie Clarion-Clipperton o wysokiej zawartości manganu. Zawiera geny o wysokiej odporności na mangan, który uważa się za adaptację do Morskiego środowiska osadowego . Zawartość metali ciężkich w obszarze C3 jest stosunkowo wysoka. Może się zdarzyć, że środowisko pokładu węgla może skutecznie indukować ekspresję genów oporności, które mogą mieć antagonistyczne i detoksykujące działanie na transport i toksyczność metali ciężkich w mikroorganizmach. Staphylococcus w tym obszarze może zawierać geny oporności przystosowujące się do środowiska pokładu węgla, dzięki czemu może również uczestniczyć w metabolizmie fermentacji węgla. Istnieje wiele różnych gatunków metanogenów w C7, a bakterie produkujące wodór są zdominowane przez Clostridium, Bacillus, Citrobacter i inne beztlenowce, które dostarczają substraty dla acetoklastycznych metanogenów, a także H2, CO2 i mrówczan dla hydrogenotropowych metanogenów. Ponadto akumulujący kwas octowy redukuje bakterie redukujące siarczany, w tym Desulfosporosinus i Desulfitobacterium. SRB mają silniejsze powinowactwo do kwasu octowego niż metanogeny acetoklastyczne, ale nie konkurują z metanogen metylotroficznych o pewne substraty, takie jak metanol. Tak więc metabolizm zarówno bakterii redukujących siarczany, jak i metanogenów może przebiegać jednocześnie w tym obszarze . Metabolizm w C7 był zdominowany przez metanogeny metylotroficzne, a następnie fermentację kwasu octowego, a następnie redukcję dwutlenku węgla.