DISCUSSÃO
determinou-se aqui o genoma tamanhos, de 25 de chironomid espécies; estes valores foram congruentes com o genoma tamanhos das três espécies anteriormente reportedin literatura (Petitpierre, 1996; Schmidt-Ott et al., 2009), que foram compostas entre 0,13 e 0,21 pg DNA. Os resultados deste estudo mostraram que os tamanhos do genoma dos quironomídeos eram uniformemente muito pequenos(Fig. 3, Quadro 1). O valor C médio obtido para a família Chironomidae é 0.13 + 0.03 PG DNA, que é claramente menor que o tamanho do genoma de D. melanogaster. O menor tamanho do genoma entre os insetos foi registrado no Strepsiptera Caenocholax fenyesi com cerca de 0,11 pg de DNA (correspondentes a cerca de 108 Mb), apesar de Hesse voar Mayetiola processo de destruição foi sugerido também a ter uma menor tamanho do genoma de 0,09 pg (Johnston et al., 2004; Gregory, 2014). Aqui, nós relatamos, pelo menos, três espécies com menores genomas do que qualquer inseto conhecido: C. tsushimensis, Diamesa japonica e Hydrobaenus tsukubalatus, com C-valores de 0,07 pg, 0,08 pg e 0,08 pg, respectivamente (Tabela 1). Tanto quanto sabemos, C. tsushimensis (Fig. 1B) Pode ser considerado como tendo o menor tamanho do genoma de qualquer inseto conhecido, com um valor C de 0,07 pg de DNA, correspondendo aproximadamente a 68 Mb. Este valor está de acordo com o tamanho do genoma de uma espécie relacionada, Clunio marinus, que foi estimada uma vez para 95 Mb e, mais recentemente, 87.2 Mb (Kaiser e Heckel, 2012; Tobias S. Kaiser, comunicação pessoal).
Recently, the genome of the Antarctic midge, B. antarctica, was sequenced (Kelley et al., 2014). Os autores alegaram que o tamanho do genoma desta espécie, estimado em 89.5-105 Mb, era o menor entre todos os insetos. No entanto, mostramos aqui que este tamanho do genoma é composto dentro da gama normal de valores de quironomídeos. Os autores também sugeriram que o pequeno tamanho do genoma de B. antarctica era uma adaptação ao ambiente frio extremo. Nossos dados podem corroborar esta hipótese, já que D. japonica, uma espécie alpina da subfamília Diamesinae também apresenta um pequeno tamanho do genoma. Outro Diamesinae relacionado, O Himalaia Midge Diamesa sp. foi encontrado ativo a temperaturas tão baixas como -16°C (Kohshima, 1984). No entanto, Telmatogeton japonicus, que também é capaz de se desenvolver a temperaturas muito baixas (Danks, 1971; Sunose e Fujisawa, 1982), mostra um tamanho relativamente Grande do genoma, em comparação com a média da família chironomid. Uma influência filogenética no tamanho do genoma deve ser tida em conta aqui, uma vez que B. Antártica pertence à subfamília Orthocladiinae, que apresenta pequenos tamanhos de genoma em média. As espécies C. tsushimensis e Hydrobaenus tsukubalatus, mostrando o menor tamanho do genoma entre os chironomídeos, também pertencem à subfamília Orthocladiinae. Assim, o pequeno tamanho do genoma de B. a Antártida pode simplesmente constituir uma característica ancestral, não o resultado de uma adaptação ao frio extremo.
P. vanderplanki é a única espécie de insecto conhecida a atingir a anidrobiose num estado completamente desidratado (Fig. 1D) e recuperar após rehidratação (Hinton, 1951, 1960a; Watanabe, 2006; Cornette e Kikawada, 2011). O nosso grupo de investigação publicou recentemente uma análise comparativa das sequências genómicas do Polipedilum vanderplanki tolerante à dessecação e do polipedilum nubifer sensível à dessecação (Gusev et al., 2014). A comparação mostrou que ambas as espécies tinham tamanhos de genoma semelhantes e que apenas um conjunto limitado de conjuntos de genes duplicados estava relacionado com a anidrobiose em P. vanderplanki (Gusev et al., 2014). Os tamanhos do genoma inferidos da montagem do genoma eram de 104 Mb para P. vanderplanki e 107 Mb para P. nubifer e estes valores não diferiam marcadamente dos tamanhos do genoma medidos para estas espécies no presente estudo, que correspondia a aproximadamente 98 Mb. P. vanderplanki e P. genomas de nubifer mostraram uma baixa proporção semelhante de repetições de ADN e apenas um pequeno número de elementos transponíveis foram encontrados, em comparação com outras espécies de dipterano. Esta redução da proporção de elementos transponíveis também foi observada na midge Antártica, B. Antártica (Kelley et al., 2014). O comprimento médio do intrão também foi consideravelmente reduzido na midge Antártica e tanto em P. vanderplanki quanto em P. nubifer (Gusev et al., 2014). Desde o sensível à dessecação e intolerante ao congelamento P. nubifer ações estas características com a Antártida, midge e o anhydrobiotic midge, uma baixa proporção de transposable elements e curto intron comprimento pode constituir uma característica compartilhada por todos os chironomids e podem não ser diretamente relacionados com a tolerância a ambientes extremos, até mesmo se adaptativo efeito não pode ser excluído.
Fig. 3.
Phylogeny of Diptera showing estimated C-values. A família Chironomid é mostrada em preto. A linha de traço mostra o tamanho do genoma de D. melanogaster. O cladograma filogenético é um consenso de diferentes árvores Quironomidas e Dipteranas (Yeates e Wiegmann, 1999, 2005; Saether, 2000; Cranston et al., 2012). As barras de erros mostram o desvio-padrão. Os tamanhos do genoma sem barras de erro foram obtidos a partir de estudos anteriores listados na Base de dados de tamanho do genoma Animal (Gregory, 2014).
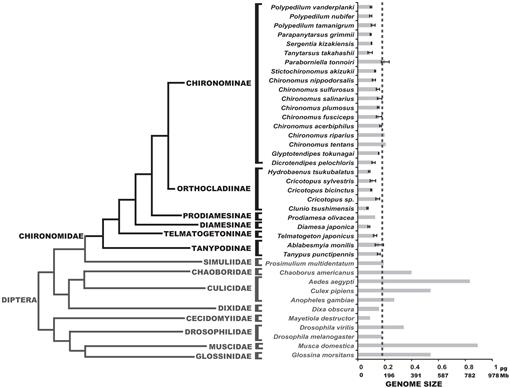
Fig. 4.
relação entre o tamanho do corpo e o tamanho do genoma entre as espécies chironómicas investigadas no presente estudo. A) para cada espécie, o comprimento total médio do corpo é indicado em milímetros e o valor C médio é expresso em pg de ADN. Somente uma correlação positiva fraca (r2 = 0.171, n = 25), materializada por uma linha sólida, foi observada entre as duas variáveis. B) ao nível do género, os meios dos mesmos valores foram transformados pelo método dos contrastes filogeneticamente independentes, com base na filogenia de Cranston et al. (2012). A correlação entre estes contrastes transformados, materializados por uma linha sólida, não foi significativa (r2 = 0.118, n = 13). A análise de dados foi realizada com Prism 4.0 software para Macintosh (GraphPad Software, Inc.).

quais poderiam ser as características adaptativas potencialmente associadas com os pequenos tamanhos do genoma entre a família Chironomidae? Na verdade, vários fatores de desenvolvimento e ecológicos têm sido sugeridos para correlacionar-se com o tamanho do genoma. Em primeiro lugar, uma correlação positiva entre o tamanho do genoma e o tamanho do corpo foi encontrada em muitos táxons e esta correlação aparece como um fenômeno relativamente geral (Gregory, 2005). Esta correlação tem sido relatada também em mosquitos (Ferrari e Rai, 1989), mas nem sempre é clara em outros táxons de insetos (Gregory e Hebert, 2003; Ardila-Garcia e Gregory, 2009). Dentro dos quironomídeos, encontramos apenas uma fraca correlação positiva entre o tamanho do genoma e o tamanho do corpo (Fig. 4A). O valor C médio para o gênero Chironomus (0,15 pg) era maior do que para o gênero Polypedilum (0,10 pg), E esta diferença pode ser atribuída ao tamanho corporal geralmente maior das espécies Chironomus. No entanto, a distância filogenética entre ambos os gêneros também poderia explicar esta diferença de tamanho do genoma. Consequentemente, corrigimos os nossos dados ao nível do género com análise de contrastes filogeneticamente independentes (Garland and Adolph, 1994; Garland et al., 2005) e como resultado, a correlação entre o tamanho do genoma e o tamanho do corpo revelou-se não significativa (Fig. 4B). A estreita gama de tamanhos do genoma Chironomídeo (apenas duas—três vezes) e o tamanho médio pequeno do corpo destas espécies (a maioria não excede alguns milímetros) provavelmente tornam difícil encontrar qualquer correlação entre ambos os traços dentro da família Chironomidae.Entre os insetos, outra correlação foi sugerida entre o tamanho do genoma e a complexidade do desenvolvimento. Enquanto os insetos hemimetabolosos, com apenas moldes ninfais graduais, apresentam uma grande variedade de tamanhos de genoma (valores C de 0,18 a 16.93 pg), insetos holometabolosos com sua metamorfose complexa têm valores C restritos dentro de um limiar putativo 2 pg, para a maioria deles (Gregory, 2002, 2005). A influência da complexidade do desenvolvimento no tamanho do genoma é aqui óbvia porque os insetos holometabolosos passam por uma remodelação morfológica intensiva durante o tempo limitado de metamorfose e um pequeno tamanho do genoma apresenta vantagens para a realização de divisões celulares a uma taxa elevada. Os chironomídeos são insectos holometabolosos e a sua metamorfose é extremamente rápida. Sua fase pupal pode durar apenas um dia, ou mesmo apenas algumas horas para certas espécies (Cranston, 1995c). Por exemplo, C. tsushimensis mostra uma metamorfose complexa com dimorfismo sexual pronunciado (Fig. 1B).
a relação entre o tamanho do genoma e a taxa de divisão celular também influencia a taxa de desenvolvimento. Por exemplo, sugere-se que o ciclo de vida rápido dos pulgões esteja relacionado com os seus pequenos tamanhos de genoma (Ma et al., 1992). Assim, as pequenas dimensões do genoma devem constituir uma vantagem para as espécies de quironomídeos que se desenvolvem rapidamente em ambientes temporários. Os chironomídeos do gênero Clunio podem se desenvolver em piscinas temporárias de marés e também mostrar o surgimento sincronizado dos adultos em relação ao ciclo lunar e durante a curta janela da maré baixa (Kaiser e Heckel, 2012). Para o conseguir, deve ser necessária uma elevada taxa de desenvolvimento. Note que os pequenos anelídeos policaetas que se desenvolvem rapidamente em ambientes intersticiais semelhantes apresentam tamanhos menores do genoma do que as espécies macrobenticas (Gambi et al., 1997). Para insetos Antárticos, como B. antarctica ou Eretmoptera murphyi (Lee et al., 2006; Worland, 2010), o desenvolvimento larval dura dois anos, na maioria presos em estado congelado, mas a metamorfose e a reprodução devem ocorrer durante o curto período de Verão. Neste caso, uma alta taxa de desenvolvimento é necessária e isso pode ser facilitado por B. antarctica pequeno genoma. Este fenômeno foi ilustrado por um estudo sobre plantas angiospermas, mostrando que as espécies capazes de completar todo o seu ciclo de vida durante o curto verão Antártico apresentaram os menores tamanhos de genoma (Bennett et al., 1982). Outro extremo, o midge anidrobiótico P. vanderplanki é adaptado para piscinas de rochas efêmeras. Mais uma vez, a elevada taxa de desenvolvimento num habitat temporário pode estar ligada ao pequeno tamanho do genoma desta espécie. No entanto, a espécie Australiana P. tonnoiri, que também está exposta à dessecação em piscinas rochosas similares (Jones, 1975; Adams, 1985; Frouz et al., 2003), mostra um tamanho relativamente Grande do genoma para um quironomide (Fig. 3 e Quadro 1).
tensões abióticas são também susceptíveis de influenciar a integridade do ADN e, como consequência, o tamanho do genoma. Por exemplo, larvas de P. vanderpianki experimentou danos massivos no DNA após a anidrobiose e a reparação eficiente do DNA ocorre durante os poucos dias seguintes à rehidratação (Gusev et al., 2010). Em plantas, uma correlação negativa foi encontrada entre o tamanho do genoma e a tolerância à radiação (Bennett e Leitch, 2005) e a taxa de mutação também foi menor em espécies com menor tamanho do genoma (Abrahamson et al., 1973). Consequentemente, o pequeno tamanho do genoma de P. vanderpianki pode representar uma vantagem para evitar a acumulação de mutações deletérias durante os eventos de reparação do ADN associados a ciclos de dessecação e reidratação. Como espécie marinha, a C. tsushimensis também está exposta ao alto estresse de salinidade, que é letal para a maioria das espécies de insetos. A alta salinidade e outras tensões abióticas são conhecidas por gerar espécies de oxigênio reativo intracelular( ROS), que foram sugeridas como uma fonte principal de dano ao DNA (Franca et al., 2007; Gill and Tuteja, 2010). Isto também pode estar relacionado ao tamanho pequeno do genoma. No entanto, outras espécies marinhas e tolerantes à salinidade elevada, como T. japonicus ou Chironomus salinarius, não apresentam tamanhos de genoma tão pequenos como C. tsushimensis. Espécies tolerantes ao ácido, tais como Chironomus sulfurosus, Chironomus acerbiphilus ou Polypedilum tamanigrum (Fig. 1C) pode sobreviver em fontes termais com um pH tão baixo quanto 1,4 (Doi et al., 2004; Takagi et al., 2005). Mais uma vez, não foi observada nenhuma correlação clara entre a tolerância ao stress ácido e o pequeno tamanho do genoma e a influência filogenética pareceu mais importante, com menor tamanho do genoma para o género Polypedilum e maiores tamanhos do genoma no género Chironomus.
para concluir, os pequenos tamanhos de genoma observados em quironomídeos são provavelmente o resultado de uma redução ancestral do número de elementos transponíveis e do comprimento dos intrões na sua estrutura genómica. Entre a família Chironomidae, os tamanhos do genoma diferiam de acordo com a filogenia no nível subfamília e no nível do gênero. Como inferido a partir de correlações em vários taxa, o pequeno tamanho do genoma dos quironomídeos teoricamente poderia ter constituído uma pré-adaptação para ambientes instáveis e extremos através de alta taxa de desenvolvimento e baixa taxa de mutação. Esperamos que este estudo encoraje futuros projetos de genoma Chironomide e que a genômica comparativa dentro deste táxon ajude a entender como os genomas Chironomides estão interagindo com seu ambiente.