discuție
am determinat aici dimensiunile genomului a 25 de specii de chironomide; aceste valori au fost congruente cu dimensiunile genomului celor trei specii raportate anterior în literatura de specialitate (Petitpierre, 1996; Schmidt-Ott și colab., 2009), care au fost cuprinse între 0,13 și 0,21 PG ADN. Rezultatele acestui studiu au arătat că dimensiunile genomului chironomidelor au fost uniform foarte mici (Fig. 3, Tabelul 1). Valoarea medie C obținută pentru familia Chironomidae este de 0,13 + 0.03 PG ADN, care este în mod clar mai mică decât dimensiunea genomului D. melanogaster. Cea mai mică dimensiune a genomului dintre insecte a fost înregistrată în Strepsiptera caenocholax fenyesi cu aproximativ 0,11 PG ADN (corespunzând aproximativ la 108 Mb), deși zbura hessiană mayetiola destructor s-a sugerat, de asemenea, să aibă o dimensiune a genomului chiar mai mică de 0,09 pg (Johnston și colab., 2004; Gregory, 2014). Aici, raportăm cel puțin trei specii cu genomi mai mici decât orice insectă cunoscută: C. tsushimensis, Diamesa japonica și Hydrobaenus tsukubalatus, cu valori C de 0,07 pg, 0,08 pg și, respectiv, 0,08 pg (Tabelul 1). După cunoștințele noastre, C. tsushimensis (Fig. 1B) poate fi considerat acum ca având cea mai mică dimensiune a genomului oricărei insecte cunoscute, cu o valoare C de 0,07 PG ADN, corespunzând aproximativ la 68 Mb. Această valoare este congruentă cu dimensiunea genomului unei specii înrudite, Clunio marinus, care a fost estimată o dată la 95 Mb și mai recent la 87,2 Mb (Kaiser și Heckel, 2012; Tobias S. Kaiser, comunicare personală).
recent, genomul musculiței Antarctice, B. antarctica, a fost secvențiat (Kelley și colab., 2014). Autorii au susținut că dimensiunea genomului acestei specii, estimată la 89.5-105 Mb, a fost cel mai mic dintre toate insectele. Cu toate acestea, arătăm aici că această dimensiune a genomului este cuprinsă în intervalul normal al valorilor chironomidelor. Autorii au sugerat, de asemenea, că dimensiunea mică a genomului B. antarctica a fost o adaptare la mediul extrem de rece. Datele noastre pot corobora această ipoteză, deoarece D. japonica, o specie alpină din subfamilia Diamesinae prezintă, de asemenea, o dimensiune mică a genomului. Un alt diamesinae înrudit, Himalaya midge Diamesa sp. s-a constatat de fapt că este activ la temperaturi de până la -16 ct (kohshima, 1984). Cu toate acestea, Telmatogeton japonicus, care este, de asemenea, capabil să se dezvolte la temperaturi foarte scăzute (Danks, 1971; Sunose și Fujisawa, 1982), prezintă o dimensiune relativ mare a genomului, comparativ cu media familiei chironomide. O influență filogenetică asupra dimensiunii genomului ar trebui luată în considerare aici, deoarece B. antarctica aparține subfamiliei Orthocladiinae, care prezintă în medie dimensiuni mici ale genomului. Speciile C. tsushimensis și Hydrobaenus tsukubalatus, care prezintă cele mai mici dimensiuni ale genomului dintre chironomide, aparțin, de asemenea, subfamiliei Orthocladiinae. Astfel, dimensiunea mică a genomului B. antarctica poate constitui pur și simplu o trăsătură ancestrală, nu rezultatul adaptării extreme la frig.
P. vanderplanki este singura specie de insecte cunoscută pentru a obține anhidrobioză într-o stare complet deshidratată (Fig. 1D) și se recuperează după rehidratare (Hinton, 1951, 1960a; Watanabe, 2006; Cornette și Kikawada, 2011). Grupul nostru de cercetare a publicat recent o analiză comparativă a secvențelor genomului Polypedilum vanderplanki tolerant la deshidratare și Polypedilum congeneric nubifer sensibil la deshidratare (Gusev și colab., 2014). Comparația a arătat că ambele specii aveau dimensiuni similare ale genomului și că doar un set limitat de grupuri de gene duplicate a fost legat de anhidrobioză la P. vanderplanki (Gusev și colab., 2014). Dimensiunile genomului deduse din asamblarea genomului au fost de 104 Mb pentru P. vanderplanki și 107 Mb pentru P. nubifer și aceste valori nu au diferit semnificativ de dimensiunile genomului măsurate pentru aceste specii în studiul de față, care au corespuns la aproximativ 98 Mb. P. vanderplanki și P. genomii nubifer au arătat o proporție similară scăzută de repetări ale ADN-ului și s-au găsit doar un număr mic de elemente transpozabile, în comparație cu alte specii dipterane. Această reducere a proporției elementelor transpozabile a fost observată și în mușchiul Antarctic, B. antarctica (Kelley și colab., 2014). Lungimea medie a intronului a fost, de asemenea, redusă considerabil în mușchiul Antarctic și atât în P. vanderplanki, cât și în P. nubifer (Gusev și colab., 2014). Deoarece uscarea sensibile și congela intolerant P. nubifer împărtășește aceste caracteristici cu mușchiul Antarctic și mușchiul anhidrobiotic, o proporție scăzută de elemente transpozabile și lungimea scurtă a intronului pot constitui o caracteristică împărtășită de toate chironomidele și pot să nu fie direct legate de toleranța la medii extreme, chiar dacă nu ar putea fi exclus un efect adaptiv.
Fig. 3.
filogenia Dipterelor care prezintă valori c estimate. Familia Chironomidă este prezentată în negru. Linia de bord arată dimensiunea genomului lui D. melanogaster. Cladograma filogenetică este un consens din diferite Chironomid și Dipteran copaci (Yeates și Wiegmann, 1999, 2005; Saether, 2000; Cranston și colab., 2012). Erori bare arată deviația standard. Dimensiunile genomului fără bare de eroare au fost obținute din studiile anterioare enumerate în baza de date Animal Genome Size (Gregory, 2014).
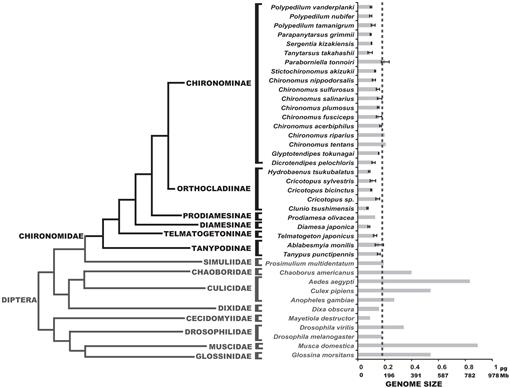
Fig. 4.
relația dintre dimensiunea corpului și dimensiunea genomului în rândul speciilor chironomide investigate în studiul de față. (A) pentru fiecare specie, lungimea medie totală a corpului este indicată în milimetri, iar valoarea medie C este exprimată în ADN-ul pg. Doar o corelație pozitivă slabă (r2 = 0,171, n = 25), materializată printr-o linie solidă, a fost observată între cele două variabile. (B) la nivel de gen, mijloacele cu aceleași valori au fost transformate prin metoda contrastelor independente filogenetic, bazate pe filogenia lui Cranston și colab. (2012). Corelația dintre aceste contraste transformate, materializate printr-o linie solidă, nu a fost semnificativă (r2 = 0,118, n = 13). Analiza datelor a fost efectuată pe Prism 4.0 software pentru Macintosh (GraphPad Software, Inc.).

care ar putea fi trăsăturile adaptive potențial asociate cu dimensiunile mici ale genomului din familia Chironomidae? De fapt, s-a sugerat că mai mulți factori de dezvoltare și ecologici se corelează cu dimensiunile genomului. În primul rând, o corelație pozitivă între dimensiunea genomului și dimensiunea corpului a fost găsită la mulți taxoni și această corelație apare ca un fenomen relativ general (Gregory, 2005). Această corelație a fost raportată și la țânțari (Ferrari și Rai, 1989), dar nu este întotdeauna clară la alți taxoni de insecte (Gregory și Hebert, 2003; Ardila-Garcia și Gregory, 2009). În cadrul chironomidelor, am găsit doar o corelație pozitivă slabă între dimensiunea genomului și dimensiunea corpului (Fig. 4A). Valoarea medie C pentru genul Chironomus (0,15 pg) a fost mai mare decât cea pentru genul Polypedilum (0,10 pg), iar această diferență poate fi atribuită dimensiunii generale mai mari a corpului speciei Chironomus. Cu toate acestea, distanța filogenetică dintre ambele genuri ar putea explica, de asemenea, această diferență de dimensiuni ale genomului. În consecință, am corectat datele noastre la nivel de gen cu analiza contrastelor independente filogenetic (Garland și Adolph, 1994; Garland și colab., 2005) și, ca urmare, corelația dintre dimensiunea genomului și dimensiunea corpului s-a dovedit a fi nesemnificativă (Fig. 4B). Gama îngustă de dimensiuni ale genomului Chironomid (doar două—trei) și dimensiunea medie mică a corpului acestor specii (majoritatea nu depășesc câțiva milimetri) fac probabil dificilă găsirea unei corelații între ambele trăsături din familia Chironomidae.
printre insecte, a fost sugerată o altă corelație între dimensiunea genomului și complexitatea dezvoltării. În timp ce insectele hemimetabolice, cu numai molturi nimfale treptate, prezintă o gamă largă de dimensiuni ale genomului (valori C de la 0,18 la 16.93 pg), insectele holometabolice cu metamorfoza lor complexă au valori C restricționate într-un prag presupus de 2 pg, pentru majoritatea acestora (Gregory, 2002, 2005). Influența complexității dezvoltării asupra dimensiunii genomului este aici evidentă, deoarece insectele holometabolice suferă o remodelare morfologică intensă în timpul limitat al metamorfozei și o dimensiune mică a genomului prezintă avantaje pentru realizarea diviziunilor celulare la o rată ridicată. Chironomidele sunt insecte holometabolice, iar metamorfoza lor este extrem de rapidă. Stadiul lor pupal poate dura doar o zi, sau chiar doar câteva ore pentru anumite specii (Cranston, 1995c). De exemplu, C. tsushimensis prezintă o metamorfoză complexă cu dimorfism sexual pronunțat (Fig. 1B).
relația dintre dimensiunea genomului și rata diviziunii celulare influențează, de asemenea, rata de dezvoltare. De exemplu, s-a sugerat că ciclul de viață rapid al afidelor este legat de dimensiunile lor mici ale genomului (Ma și colab., 1992). Astfel, dimensiunile mici ale genomului ar trebui să constituie un avantaj pentru speciile chironomide care se dezvoltă rapid în medii temporare. Chironomidele din genul Clunio se pot dezvolta în bazine temporare de maree și prezintă, de asemenea, apariția sincronizată a adulților în raport cu ciclul Lunii și în timpul ferestrei scurte a mareei joase (Kaiser și Heckel, 2012). Ar trebui să fie necesară o rată ridicată de dezvoltare pentru a realiza acest lucru. Rețineți că anelidele Polichete mici care se dezvoltă rapid în medii interstițiale similare prezintă dimensiuni mai mici ale genomului decât speciile macrobentice (Gambi și colab., 1997). Pentru musculițele Antarctice, cum ar fi B. antarctica sau Eretmoptera murphyi (Lee și colab., 2006; Worland, 2010), dezvoltarea larvelor durează doi ani, cea mai mare parte arestată într-o stare înghețată, dar metamorfoza și reproducerea trebuie să aibă loc în perioada foarte scurtă de vară. În acest caz, este necesară o rată ridicată de dezvoltare și acest lucru poate fi facilitat de genomul mic B. antarctica. Acest fenomen a fost ilustrat de un studiu asupra plantelor angiosperme, arătând că speciile capabile să-și finalizeze întregul ciclu de viață în timpul verii Antarctice scurte au prezentat cele mai mici dimensiuni ale genomului (Bennett și colab., 1982). O altă extremă, mușchiul anhidrobiotic P. vanderplanki este adaptat la bazinele de rocă efemere. Din nou, rata mare de dezvoltare într-un habitat temporar poate fi legată de dimensiunea mică a genomului acestei specii. Cu toate acestea, specia australiană P. tonnoiri, care este, de asemenea, expusă deshidratării pe bazine de rocă similare (Jones, 1975; Adams, 1985; Frouz și colab., 2003), prezintă o dimensiune relativ mare a genomului pentru un chironomid (Fig. 3 și Tabelul 1).
tensiunile abiotice sunt, de asemenea, susceptibile de a influența integritatea ADN-ului și, în consecință, dimensiunea genomului. De exemplu, larvele de P. vanderpianki prezintă leziuni masive ale ADN – ului după anhidrobioză și repararea eficientă a ADN-ului are loc în câteva zile după rehidratare (Gusev și colab., 2010). La plante, s-a găsit o corelație negativă între dimensiunea genomului și toleranța la radiații (Bennett și Leitch, 2005), iar rata mutației a fost, de asemenea, mai mică la speciile cu dimensiunea genomului mai mică (Abrahamson și colab., 1973). În consecință, dimensiunea mică a genomului P. vanderpianki poate reprezenta un avantaj pentru a evita acumularea de mutații dăunătoare în timpul evenimentelor de reparare a ADN-ului asociate ciclurilor de deshidratare și rehidratare. Ca specie marină, C. tsushimensis este, de asemenea, expus la stres ridicat de salinitate, care este letal pentru majoritatea speciilor de insecte. Salinitatea ridicată și alte stresuri abiotice sunt cunoscute pentru a genera specii de oxigen reactiv intracelular (ros), care au fost sugerate ca o sursă principală de deteriorare a ADN-ului (Franca și colab., 2007; Gill și Tuteja, 2010). Acest lucru ar putea fi, de asemenea, legat de dimensiunea mică a genomului. Cu toate acestea, alte specii marine și tolerante la salinitate ridicată, cum ar fi T. japonicus sau Chironomus salinarius nu prezintă dimensiuni ale genomului la fel de mici ca C. tsushimensis. Specii tolerante la Acid, cum ar fi Chironomus sulfurosus, Chironomus acerbiphilus sau Polypedilum tamanigrum (Fig. 1C) poate supraviețui în izvoarele termale cu un pH de până la 1,4 (Doi și colab., 2004; Takagi și colab., 2005). Din nou, nu s-a putut observa o corelație clară între toleranța la stresul acid și dimensiunea mică a genomului, iar influența filogenetică a apărut mai importantă, cu o dimensiune mai mică a genomului pentru genul Polypedilum și dimensiuni mai mari ale genomului în genul Chironomus.
pentru a concluziona, dimensiunile mici ale genomului observate la chironomide sunt probabil rezultatul unei reduceri ancestrale a numărului de elemente transpozabile și a lungimii intronilor în structura genomului lor. Dintre familia Chironomidae, dimensiunile genomului diferă în funcție de filogenie la nivel de subfamilie și la nivel de gen. După cum se deduce din corelațiile din diferiți taxoni, dimensiunea mică a genomului chironomidelor teoretic ar fi putut constitui o preadaptare la medii instabile și extreme prin rata ridicată de dezvoltare și rata scăzută de mutație. Sperăm că acest studiu va încuraja viitoarele proiecte de genom Chironomid și că genomica comparativă din cadrul acestui taxon va ajuta la înțelegerea modului în care genomii Chironomid interacționează cu mediul lor.