diskussion
vi bestämde här genomstorlekarna för 25 chironomidarter; dessa värden var kongruenta med genomstorlekarna för de tre arterna som tidigare rapporterats i litteraturen (Petitpierre, 1996; Schmidt-Ott et al., 2009), som bestod mellan 0,13 och 0,21 pg DNA. Resultaten av denna studie visade att genomstorlekarna hos chironomider var enhetligt mycket små (Fig. 3, Tabell 1). Det erhållna medelvärdet C – värdet för familjen Chironomidae är 0,13 + 0.03 pg DNA, vilket är klart mindre än D. melanogaster genomstorlek. Den minsta genomstorleken bland insekter har registrerats i Strepsiptera Caenocholax fenyesi med cirka 0,11 pg DNA (motsvarande ungefär 108 Mb), även om den hessiska flugan Mayetiola destructor också föreslogs ha en ännu mindre genomstorlek på 0,09 pg (Johnston et al., 2004; Gregory, 2014). Här rapporterar vi minst tre arter med mindre genom än någon känd insekt: C. tsushimensis, Diamesa japonica och Hydrobaenus tsukubalatus, med C-värden på 0,07 pg, 0,08 pg respektive 0,08 pg (Tabell 1). Så vitt vi vet, C. tsushimensis (Fig. 1B) kan nu anses ha den minsta genomstorleken av någon känd insekt, med ett C-värde på 0,07 pg DNA, motsvarande ungefär 68 Mb. Detta värde överensstämmer med genomstorleken hos en besläktad Art, Clunio marinus, som uppskattades en gång till 95 Mb och mer nyligen till 87, 2 Mb (Kaiser och Heckel, 2012; Tobias S. Kaiser, personlig kommunikation).
nyligen sekvenserades genomet av Antarktis midge, B. Antarktis (Kelley et al., 2014). Författarna hävdade att genomstorleken hos denna art, uppskattad till 89.5-105 Mb, var den minsta bland alla insekter. Men vi visar här att denna genomstorlek består av det normala intervallet av chironomidsvärden. Författarna föreslog också att den lilla genomstorleken hos B. Antarktis var en anpassning till extrem kall miljö. Våra data kan bekräfta denna hypotes, eftersom D. japonica, en alpin art från underfamiljen Diamesinae presenterar också en liten genomstorlek. En annan relaterad Diamesinae, Himalaya midge Diamesa sp. var faktiskt befunnits vara aktiv vid temperaturer så låga som -16 kcal C (Kohshima, 1984). Telmatogeton japonicus, som också kan utvecklas vid mycket låga temperaturer (Danks, 1971; Sunose och Fujisawa, 1982), visar emellertid en relativt stor genomstorlek jämfört med chironomid-familjen. Ett fylogenetiskt inflytande på genomstorleken bör beaktas här, eftersom B. Antarktis tillhör Orthocladiinae-underfamiljen, som i genomsnitt presenterar små genomstorlekar. Arten C. tsushimensis och Hydrobaenus tsukubalatus, som visar de minsta genomstorlekarna bland chironomider, tillhör också Orthocladiinae-underfamiljen. Således är den lilla genomstorleken av B. Antarktis kan helt enkelt utgöra ett förfäderdrag, inte resultatet av extrem kall anpassning.
P. vanderplanki är den enda insektsarten som är känd för att uppnå anhydrobios i ett helt uttorkat tillstånd (Fig. 1D) och återhämta sig efter rehydrering (Hinton, 1951, 1960a; Watanabe, 2006; Cornette och Kikawada, 2011). Vår forskargrupp publicerade nyligen en jämförande analys av genomsekvenserna av den torkningstoleranta Polypedilum vanderplanki och den torkningskänsliga kongeneriska Polypedilum nubifer (Gusev et al., 2014). Jämförelsen visade att båda arterna hade liknande genomstorlekar och att endast en begränsad uppsättning duplicerade genkluster var relaterade till anhydrobios I P. vanderplanki (Gusev et al., 2014). Genomstorlekarna som härleddes från genomaggregatet var 104 Mb för P. vanderplanki och 107 Mb för P. nubifer och dessa värden skilde sig inte markant från genomstorlekarna uppmätta för dessa arter i föreliggande studie, vilket motsvarade ungefär 98 Mb. P. vanderplanki och P. nubifer genom visade en liknande låg andel DNA-upprepningar och endast ett litet antal transponerbara element hittades jämfört med andra dipteranarter. Denna minskning av andelen transponerbara element observerades också i Antarktis midge, B. Antarktis (Kelley et al., 2014). Den genomsnittliga intronlängden minskades också avsevärt i Antarktis midge och i både P. vanderplanki och P. nubifer också (Gusev et al., 2014). Eftersom uttorkning känslig och frysa intolerant P. nubifer delar dessa funktioner med Antarktis midge och anhydrobiotic midge, en låg andel transponerbara element och kort intron längd kan utgöra en egenskap som delas av alla chironomider och kan inte vara direkt relaterad till toleransen mot extrema miljöer, även om en adaptiv effekt inte kunde uteslutas.
Fig. 3.
fylogeni av Diptera som visar uppskattade C-värden. Familjen Chironomid visas i svart. Strecklinjen visar genomstorleken på D. melanogaster. Det fylogenetiska kladogrammet är en konsensus från olika Chironomid-och Dipteranträd (Yeates och Wiegmann, 1999, 2005; Saether, 2000; Cranston et al., 2012). Fel barer visar standardavvikelse. Genomstorlekar utan felstänger erhölls från tidigare studier listade i databasen för Djurgenomstorlek (Gregory, 2014).
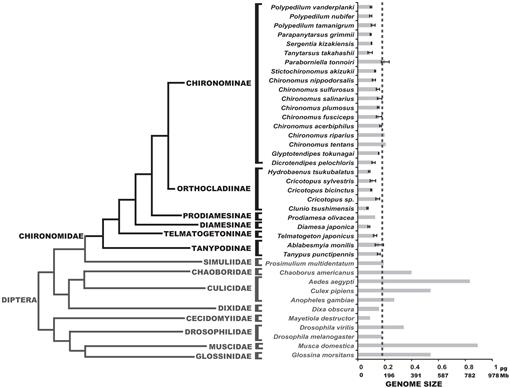
Fig. 4.
förhållandet mellan kroppsstorlek och genomstorlek bland de chironomidarter som undersöktes i den aktuella studien. A) för varje art anges den genomsnittliga totala kroppslängden i millimeter och medelvärdet C-värdet uttrycks i pg-DNA. Endast en svag positiv korrelation (r2 = 0,171, n = 25), materialiserad av en solid linje, observerades mellan de två variablerna. (B) på genusnivå transformerades medel för samma värden med metoden för fylogenetiskt oberoende kontraster, baserat på fylogeni av Cranston et al. (2012). Korrelationen mellan dessa transformerade kontraster, materialiserad av en solid linje, var inte signifikant (r2 = 0,118, n = 13). Dataanalys utfördes på Prism 4.0 programvara för Macintosh (GraphPad Software, Inc.).

vad kan vara de adaptiva egenskaperna som potentiellt är förknippade med de små genomstorlekarna bland familjen Chironomidae? Faktum är att flera utvecklings-och ekologiska faktorer har föreslagits att korrelera med genomstorlekar. Först har en positiv korrelation mellan genomstorlek och kroppsstorlek hittats i många taxa och denna korrelation framträder som ett relativt allmänt fenomen (Gregory, 2005). Denna korrelation har rapporterats också i myggor (Ferrari och Rai, 1989), men det är inte alltid klart i andra insektstaxa (Gregory och Hebert, 2003; Ardila-Garcia och Gregory, 2009). Inom chironomiderna fann vi bara en svag positiv korrelation mellan genomstorlek och kroppsstorlek (Fig. 4A). Medelvärdet C-värdet för släktet Chironomus (0,15 pg) var högre än för släktet Polypedilum (0,10 pg), och denna skillnad kan hänföras till den generellt större kroppsstorleken hos Chironomus-arter. Det fylogenetiska avståndet mellan båda släktena kan emellertid också förklara denna skillnad i genomstorlekar. Följaktligen korrigerade vi våra data på genusnivå med fylogenetiskt oberoende kontrastanalys (Garland och Adolph, 1994; Garland et al., 2005) och som ett resultat visade sig korrelationen mellan genomstorlek och kroppsstorlek vara icke signifikant (Fig. 4B). Det smala området av Chironomidgenomstorlekar (endast två—trefaldigt) och den genomsnittliga lilla kroppsstorleken hos dessa arter (de flesta överstiger inte några millimeter) gör det förmodligen svårt att hitta någon korrelation mellan båda egenskaperna inom Chironomidae-familjen.
bland insekter föreslogs en annan korrelation mellan genomstorlek och utvecklingskomplexitet. Medan hemimetabolous insekter, med gradvisa nymphal molts endast, visar ett brett spektrum av genomstorlekar (C-värden från 0,18 till 16.93 pg), holometabolous insekter med deras komplexa metamorfos har C-värden begränsade inom en förmodad 2 pg tröskel, för de flesta av dem (Gregory, 2002, 2005). Påverkan av utvecklingskomplexitet på genomstorlek är här uppenbart eftersom holometabolous insekter genomgår intensiv morfologisk ombyggnad under den begränsade tiden av metamorfos och en liten genomstorlek ger fördelar för att åstadkomma celldelningar i hög takt. Chironomider är holometabolous insekter och deras metamorfos är extremt snabb. Deras pupalstadium kan bara vara en dag, eller till och med bara några timmar för vissa arter (Cranston, 1995c). Till exempel visar C. tsushimensis en komplex metamorfos med uttalad sexuell dimorfism (Fig. 1B).
förhållandet mellan genomstorlek och graden av celldelning påverkar också utvecklingshastigheten. Till exempel föreslogs den snabba livscykeln för bladlöss att vara relaterad till deras små genomstorlekar (Ma et al., 1992). Således bör små genomstorlekar utgöra en fördel för chironomidarter som snabbt utvecklas i tillfälliga miljöer. Chironomider från släktet Clunio kan utvecklas till tillfälliga tidvattenpooler och visar också synkroniserad uppkomst av de vuxna i förhållande till måncykeln och under lågvattenets korta fönster (Kaiser och Heckel, 2012). En hög utvecklingsgrad bör behövas för att uppnå detta. Observera att små polychaete annelider utvecklas snabbt i liknande interstitiella miljöer visar mindre genomstorlekar än makrobenthic arter (Gambi et al., 1997). För Antarktiska midger, såsom B. Antarktis eller Eretmoptera murphyi (Lee et al., 2006; Worland, 2010), larvutveckling varar i två år, mestadels arresterad i fryst tillstånd, men metamorfos och reproduktion måste ske under den mycket korta sommarperioden. I detta fall behövs en hög utvecklingshastighet och detta kan underlättas av B. Antarktis lilla genom. Detta fenomen illustrerades av en studie på angiospermväxter som visade att arten som kunde slutföra hela sin livscykel under den korta Antarktiska sommaren presenterade de minsta genomstorlekarna (Bennett et al., 1982). En annan extrem, den anhydrobiotiska midge P. vanderplanki är anpassad till efemära bergpooler. Även här kan hög utvecklingshastighet i en tillfällig livsmiljö kopplas till den lilla genomstorleken hos denna art. Men den australiensiska arten P. tonnoiri, som också utsätts för uttorkning på liknande bergpooler (Jones, 1975; Adams, 1985; Frouz et al., 2003), visar en relativt stor genomstorlek för en chironomid (Fig. 3 och Tabell 1).
abiotiska påfrestningar påverkar också DNA-integritet och som en konsekvens genomstorlek. Till exempel larver av P. vanderpianki upplever massiv DNA-skada efter anhydrobios och effektiv DNA-reparation sker under de få dagarna efter rehydrering (Gusev et al., 2010). I växter hittades en negativ korrelation mellan genomstorlek och strålningstolerans (Bennett och Leitch, 2005) och mutationshastigheten var också lägre hos arter med mindre genomstorlek (Abrahamson et al., 1973). Följaktligen kan den lilla genomstorleken hos P. vanderpianki representera en fördel för att undvika ackumulering av skadliga mutationer under DNA-reparationshändelserna associerade med cykler av uttorkning och rehydrering. Som en marin Art utsätts C. tsushimensis också för hög salthalt, vilket är dödligt för de flesta insektsarter. Hög salthalt och andra abiotiska påkänningar är kända för att generera intracellulära reaktiva syrearter (ROS), som föreslogs som en huvudkälla till DNA-skada (Franca et al., 2007; Gill och Tuteja, 2010). Detta kan också vara relaterat till liten genomstorlek. Men andra marina och hög salthalt toleranta arter, såsom T. japonicus eller Chironomus salinarius visar inte genomstorlekar så små som C. tsushimensis. Syratoleranta arter, såsom Chironomus sulfurosus, Chironomus acerbiphilus eller Polypedilum tamanigrum (Fig. 1C) kan överleva i varma källor med ett pH så lågt som 1,4 (Doi et al., 2004; Takagi et al., 2005). Här igen kunde ingen tydlig korrelation mellan tolerans mot sur stress och liten genomstorlek observeras och det fylogenetiska inflytandet verkade viktigare, med mindre genomstorlek för släktet Polypedilum och större genomstorlekar i släktet Chironomus.
avslutningsvis är de små genomstorlekarna som observerats i chironomider troligen resultatet av en förfädersminskning av antalet transponerbara element och längden på introner i deras genomstruktur. Bland familjen Chironomidae skilde sig genomstorlekarna beroende på fylogeni på subfamiljenivå och på genusnivå. Som framgår av korrelationer i olika taxa kunde den lilla genomstorleken hos chironomider teoretiskt ha utgjort en preadaptation till instabila och extrema miljöer genom hög utvecklingshastighet och låg mutationshastighet. Vi hoppas att denna studie kommer att uppmuntra framtida Chironomidgenomprojekt och att jämförande genomik inom detta taxon hjälper till att förstå hur Chironomidgenom interagerar med sin miljö.