analys av faktorer som påverkar den mikrobiella samhällsstrukturen
för att ytterligare undersöka möjliga samband mellan miljöfaktorerna och samhällsvariansen skapades RDA-analys (Fig. 7). Totalt åtta miljöfaktorer, inklusive spårämnen Fe, Co, Ni, temperatur, salthalt, djup, fukt och RO. Djupet och reservoartemperaturen mättes på provtagningsplatsen och annan information erhölls från geologiska data för lokala gruvor (se Tabell 2). Data som visas i Fig. 7a avslöjade att de bakteriella samhällskompositionerna som hittades i denna studie påverkades signifikant av Fe, Ni, fukt, salthalt och RO. Alla samhällen, andra än C4, C7, C10, är positivt korrelerade med RO; C4, C7 och C10 positivt korrelerade med Fe, Ni och fukt. Co krävs för co-enzym m metyl-transferas, vilket är ett viktigt enzym i den biokemiska metabolismen av metanogener ; därför är effekten av Co på archaeal community större än den hos bakteriesamhället. Elementen Fe, Co, och Ni; liksom fukt, verkade vara de viktigaste miljöfaktorer följt av RO och salthalt i archaeal samhället. Det finns en signifikant positiv korrelation mellan Co och C1, C8, C9, C10 samhällen. Alla samhällen utom C2, C4, C6 och C5 var negativt korrelerade med salthalt (Fig. 7b).

rda (redundansanalys) baserat på nivån av bakterier (a) och archaea (b) med kolbäddens miljöfaktorer och kolegenskaper. Påverkningsfaktorns längd är längre, påverkans bidrag är högre, och omvänt, när påverkningsfaktorn är kortare, är påverkans bidrag lättare. När miljöfaktorn är akut vinklad med provet finns det en positiv korrelation, och när miljöfaktorn och provvinkeln är trubbiga finns det en negativ korrelation
Coal rank
coalification ”jump” avser en serie fysiska och kemiska förändringar under kolens temperatur och tryck under geologisk historia. Kol har således genomgått en process av gradvis till plötslig förändring. De fyra hoppen motsvarar RO på 0,6, 1.3, 2,5 och 3,0%. Oavsett archaeal eller bakteriesamhället som behandlas har kolrankningen ett visst inflytande på bakteriens mångfald och överflöd. Med en ökning av kol rang, i både archaeal och bakteriesamhällen, mångfalden i samhället visar en viss nedåtgående trend totalt sett (Fig. 8). Dessutom kan mikroorganismer påverka sammansättningen av kolet som kontrolleras av kolraderna. Mellersta och låga kol innehåller stora mängder växtevolverade ämnen i Grupp 1, som innehåller många växtevolverade ämnen. Här finns det högre innehåll av väte, syre och kväve; och de näringsämnen som krävs av bakterierna är rikliga. I processen med koalifiering genererar organiska ämnen mycket fukt och flytande kolväten. Samtidigt är sidokedjorna av väte och syre i kol också rikliga. Dessa flytande och fasta ämnen utgör grunden för livet för bakterier. Som ett resultat är överflödet och mångfalden av väteproducerande bakterier och metanogener i kol i denna region relativt höga. Med ökningen av Ro reduceras sidokedjans innehåll av väte och syre i kol drastiskt och komponenterna som är tillgängliga för mikroorganismerna reduceras också. Därför reduceras artens överflöd och mångfald i bakterie-och archaealsamhällena i Grupp 2 och Grupp 3 Totalt sett. Hittills har kolnivåerna av biogent kolbäddmetan i naturen visat sig ha en reflektivitet på 2, 0% (C4 Hebi). Efter RO > 2,5% har de organiska föreningarna som kan omvandlas till små molekyler varit mycket sällsynta, men det har varit en högre mångfald och överflöd i Grupp 4. Vi spekulerar i att näringsämnena som införs av grundvatten vid denna tidpunkt är tillgängliga för bakteriell reproduktion. De näringsämnen som grundvattnet orsakar i olika regioner och under olika årstider kan ha bidragit till mångfalden av arter. En orsak till den högre floramångfalden i Grupp 4 kan vara att C8 Jiaozuo Jiulishan-området har bättre avrinningsförhållanden för grundvatten och starkare laddning. Det kan transportera näringsämnen för floran, så mångfalden och överflödet är högre än Grupp 2. Det är värt att notera att mångfalden och överflöd av archaeal samhällen är negativt korrelerade med kolrader i viss utsträckning. Artens överflöd i bakteriesamhällena är dock positivt korrelerat med kolrader, och mångfalden visar en nedåtgående trend. Med ökningen av kolrader anpassas vissa bakteriegrupper gradvis till miljön i olika kolrader och kan växa och föröka sig i stort antal, och metanogener är svåra att anpassa sig till kolrader.
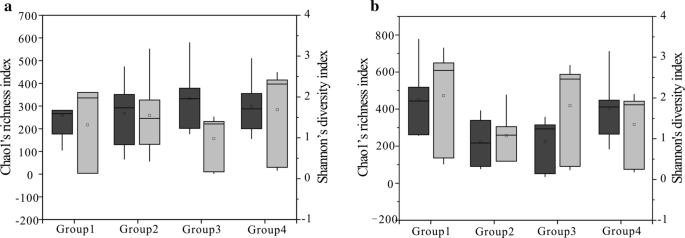
Chao1s (mörkgrå) och Shannons (ljusgrå) index för de fyra grupperna (Kolprover delades in i fyra grupper enligt värdet av RO för bakteriegemenskapen (a) och archaea-gemenskapen (b), Grupp 1 representerar ett värde mindre än 0,6%, Grupp 2 representerar värdet mellan 0,8 och 1,1%, Grupp 3 representerar värdet mellan 1,4 och 1,8%, Grupp 4 representerar värdet mellan 2,67 och 3,15%) härledda från regioner. 25: e och 75: e percentiler indikeras av lådans ytterkanter medan Max-och minimivärdena visas av ändarna på morrhåren och medianen med en horisontell linje inom varje låda
spårelement
spårelement kan främja tillväxten av mikroorganismer inom ett visst område, där cellen upprätthåller homeostas av elementen genom metabolisk reglering. Spårmetallelement kan också existera i olika enzymer, som kan absorberas och användas av mikroorganismer i processen med anaerob metabolism, vilket påverkar samhällsstrukturen hos väteproducerande bakterier och metanogener (tabell 3).
Fe och Ni har en större effekt på väteproducerande bakterier än Co . Fe och Ni kan delta i syntesen och metabolismen av hydrogenaser och andra metalloenzymer i mikroorganismer. När innehållet i Fe och Ni ökar inom ett visst intervall, så gör överflöd och mångfald av väteproducerande bakteriepopulationer. Innehållet av Fe och Ni i C7 är mycket högre än i andra regioner, och detta arbete har funnit att Clostridium är väteproducerande bakterier. Detta resultat indikerar att överdrivna nivåer av spårelementen kan ha en toxisk effekt på tillväxten av mikroorganismer och hämma aktiviteten hos metalloenzymer. Nivåerna av Fe i C4, C6 och C9 var inte signifikant olika och var stabila vid 3500 mg Kg−1 (Fig. 9). Det relativa överflödet av Ni i de tre områdena är C6 > C9 > C4, vilket motsvarar överflödesordningen (även C6 > C9 > C4), men mångfaldsordningen är C6 > C4 > C9. Medlemmar av släkten: Clostridium, Klebsiella, Enterobacter och Citrobacter upptäcktes i C4, C6 och C9 samhällen; inklusive de med högre överflöd och nivåer av mångfald än från andra regioner.

innehållet av Spårmetallelement i Fe, Co, Ni i kolprover
i archaeal-samhället är påverkan av Fe, Co och Ni på metanogenerna ännu viktigare. Co är ett nyckelelement i syntesen av metanogent koenzym F430 , och Co-innehållet i de tre bästa är C8 > C7 > C10, metanogenarter och överflöd är C8 > C7 > C10. Innehållet av Co är positivt korrelerat till överflöd och mångfald av metanogener i viss utsträckning. Även om innehållet av Fe i C7 är mycket högre än i andra regioner, påverkar det inte fördelningen av metanogener i regionen. Det finns bara få typer av metanogener som kan innehålla Fe—i tidigare studier upptäcktes endast en art, som heter Methanotermobacter. Närvaron av monoferriskt hydrogenas i metanogener av M. marburgensis katalyserar den reversibla reaktionen av metenyl-H4MPT+ och H2 för att generera metylen-H4MPT och H+; producerar metan från CO2 och H2 . Metanogener som använder hydrogenotrofisk metabolism kan också innehålla liknande enzymer. Dessutom spekuleras också en stor andel metylotrofa metanogener om att hysa sådana enzymer utom Metanokulleus och Metanobakterier. Det spekuleras att det kan finnas ett metalloenzym associerat med Fe i de metylotrofa metanogenerna.
Grundvattenförhållanden
grundvatten utgör direkt eller indirekt en ekologisk grund för tillväxt och metabolism av extremofiler i kolsömmen. Å ena sidan levererar grundvattenuppladdning stora mängder näringsämnen för bakteriella och archaeala samhällen; å andra sidan påverkar grundvattnets miljöförhållanden (Eh, pH, salthalt, jonkomposition och spårämnen) direkt mikrobiell tillväxt och metabolisk enzymaktivitet. Grundvatten miljöförhållanden är direkt relaterade till användning och nedbrytning av kol, och mikroorganismerna i kolsömmen visar olika samhällsstrukturer och funktionella egenskaper.
mikrobiella näringsämnen är i allmänhet upplösta. Avrinningszonen i gruvområdet kan möjliggöra överlevnad av kolsömmar. Reservoarer med hög permeabilitet har en positiv inverkan på tillväxten och reproduktionen av väteproducerande bakterier och metanogener, medan metamorfism har en signifikant negativ inverkan på kolpermeabiliteten i kolreservoarer . I områden med Biogen CBM har C2, C4, C6 och C7 samhällen alla dokumenterats väl. Dessa samhällen tillhör den låga och medelstora kolrankningen, kolens porositet är relativt högre än kol med hög rang, grundvatten kan ge näringsämnen till mikroberna i kolsömmen i tid. Den nuvarande CBM – utvecklingszonen inom Powder River Basin i USA. är huvudsakligen koncentrerad i avrinningszonen för grundvatten. Gasstabila isotopdata från en grund CBM-brunn i C6-gruvområdet bekräftade också förekomsten av biogenetisk CBM i området. Emellertid indikerade gasstabil isotopdata från en annan djup CBM att CBM huvudsakligen är termogen. Dessa resultat visar att när djupet av begravningen ökar kommer avrinningsförhållandena att försvagas och det blir svårt att transportera näringsämnen för mikroorganismen, vilket kommer att leda till en minskning av samhällets överflöd och mångfald. Taket och golvet i nr. 2 kolsöm i C2-området har relativt stabila lager av lersten och lersten, vilket gör det svårt för de väteproducerande bakterierna och metanogenerna i kolsömmen att erhålla flytande näringsämnen och begränsar deras tillväxt och metabolism och därmed deras mångfald och överflöd i samhället. Observera att Chao1-indexet i detta område är 240 och Shannon-indexet är 1,38 i bakteriegemenskapen. Chao1-indexet för metanogener är 82, Shannon-indexet är 0,56. Sandstenen sprickade akvifertaket i C4-området i nr. 21 kolsömmen har bättre laddningsförhållanden och fyller kolsömmen med vatten. Det är möjligt att det mikrobiella samhället upplever kumulativa effekter från tillräcklig tillgänglighet av olika näringsämnen, vilket påverkar transporten, jämfört med överflödet och mångfalden i mikroorganismsamhället i C2-området, vilket har förbättrats avsevärt. I detta område är Chao1-indexet för väteproducerande bakterier 148, Shannon-indexet är 1,52; Chao1-indexet för metanogener är 368 och Shannon-indexet är 2,35. Sandstenfissurerad akvifer av C6-området är en direkt vattenfylld akvifer av nr 3 kolsöm. Frakturutveckling inom skiktet och måttlig akveositet spelar också en aktiv roll i samhällets överflöd och mångfald. Här är Chao1-indexet för väteproducerande bakterier 472, Shannon-indexet är 1,56; Chao1-indexet för metanogener är 384 och Shannon-indexet är 1,08. Detta är också fallet i C7-området, kolsömmen nr 5 har en direkt sprickad akvifer med goda laddningsförhållanden, överflödesindexet för de väteproducerande bakterierna är 458, Shannon-indexet är 1,98; och Chao1-indexet för metanogenerna är 256 och Shannon-indexet är 2,47. Därför är artens mångfald av väteproducerande bakterier och metanogener i C4, C6 och C7 högre än i C2.
grundvatten miljöförhållanden kommer direkt att påverka tillväxten och metabolismen av mikroorganismer. PH-värdet för grundvatten i kolbädden är i allmänhet neutralt, men pH−värdet varierar mellan 6,5 och 8,4 i sandsten sprickad akvifer i C4-område nr 21 kolsöm och salthalten är 1,0 g L-1. I det direkta akviferskiktet i nr. 3 kolsöm i C6-området varierar pH mellan pH 6,8 till 8,0 och salthalten är 0,7 g L−1. Grundvattnets pH-värde för C7-området är 6,1-7,3 och salthalten är 1,25 g L−1. PH-värdet i C4, C6 och C7 är nära neutralt och graden av mineralisering är låg, där mikroorganismsamhället har bättre tillväxt, högre överflöd och högre mångfald. Dessutom är grundvattnets salthalt och jonkomposition nära besläktade med den anaeroba reduktionsmiljön i kolsömmen. Till exempel, SO42− används för att utvärdera de slutna förhållandena för grundvatten, och HCO3− är produkten av den anaeroba avsvavlingsreaktionen av SO42−, så hög HCO3− kan användas som ett tecken på god tätning och stark minskning av kolbädds grundvatten . Vattenkemin i C4-området är HCO3 * SO4-Ca * Mg, vattenkemin i C7–området liknar C4-området, HCO3·SO4-Ca·Na och ger en relativt sluten anaerob miljö. I detta fall är Chao1-indexet för väteproducerande bakterier i C4 148, Shannon-indexet är 1,52; Chao1-indexet för metanogener är 368, Shannon-indexet är 2,35. Chao1-indexet för väteproducerande bakterier i C7 är 458, Shannon-indexet är 1,98, Chao1-indexet är 256 och Shannon-indexet är 2,47. I C6 är vattenkemin SO4 * HCO3-K * Na och SO42− dominerar, medan Chao1-indexet för väteproducerande bakterier i C6 är 472, Shannon-indexet är 1,56, Chao1-indexet för metanogener är 384 och Shannon-indexet är 1,08. Data visar att mångfalden i C6 är något lägre än C4 och C7.
vissa väteproducerande bakterier och metanogener detekterades i C8-och C9-områden i det område där ingen biometan hittades. Det var också anmärkningsvärt att grundvattenförhållandena i dessa två områden liknar dem i de ovan nämnda biogena metanområdena, som ligger i grundvattenavrinningszonen och grundvattenuppladdningen är mer kapabel att transportera lite organiskt material i kolsömmen, så att ett stort antal bakterier växer och multiplicerar, vilket är en av anledningarna till den högre artens överflöd och mångfald av C8 och C9.
temperatur
temperatur och spårelement påverkar överflöd och mångfald av mikrobiella samhällen genom att direkt ändra både tillväxt och metabolism av mikroorganismer och deras metaboliska miljö. Således, från en mikrobiologisk synvinkel, är optimal temperatur en av de viktigaste faktorerna som påverkar mikroorganismernas tillväxt och metabolism. Figur 5b visar att temperaturen utövar ett relativt svagt inflytande på överflöd och mångfald av metanogener, även om väteproducerande bakterier finns inom en smal ekologisk amplitud och är känsliga för temperaturförändring. Denna variabel är korrelerad med artens överflöd och mångfald, och resultaten av denna studie visar att temperaturen på kolsömmen (dvs. mellan 25 och 27 kcal C) är positivt korrelerad med bakteriepopulationens överflöd. Vid C8 var temperaturen 27,2 CCB, den högsta temperaturen som registrerades i denna studie. Chao1-indexet för väteproducerande bakterier var 510 och Shannon–indexet var 2,61, också det högsta bland proverna (C1-C8). Den lägsta temperaturen, 24,9 CCB, hittades vid C1, där Chao1-indexet för bakteriesamhället var det lägsta. Överflödet och mångfalden av mikrobiella arter ökar med temperaturen i C3 > C6 > C2 > C7 > C5 > C4. Geotermiska gradientavvikelser vid C9 och C10 orsakade mycket högre temperaturer; omgivningstemperaturerna vid C9 och C10 var 34,60 cccrespektive 40,10 cccrespektive. Chao1-indexet för väteproducerande bakterier i C9 var 176 och Shannon-indexet var 1,28; Chao1-indexet är 237, Shannons index är 1,58. Jämfört med de första åtta områdena har överflöd och mångfald minskat något. Här kan både väteproducerande bakterier och metanogener växa och reproducera vid omgivningstemperaturen.
mikrobiella syntrofa interaktioner
i den extrema miljön i kolsömmen bildas konsortier av bakterier bland mikroorganismerna i kolsömmen. Genom utbyte av metaboliter och mikromiljöstyrd symbios upprätthåller konkurrens och resursfördelningar de specifika funktionerna i det mikrobiella samhället, som bestämmer biometanproduktionsvägen i kolsömmen. Methanothrix, som omvandlar ättiksyra till metan, är det dominerande släktet i metanogensamhället i C1-området. Bakterierna associerade med Alkalibaculum och Desulfosporosinus är homoacetogena bakterier som använder H2 som elektrondonator för att producera ättiksyra. De är huvudkonkurrenterna för hydrogenotrofa metanogener och ger också metaboliskt substrat för metanogener. Väteproducerande bakterier som Clostridium och Tissierella ger också ättiksyra, och därmed ger den höga överflöd av väteproducerande bakterier ett rikt metaboliskt substrat för Metanothrix. Tillsammans är metanogenerna och de väteproducerande bakterierna i syntrofisk interaktion och metangenereringsvägen i detta område bestäms av sönderdelning av ättiksyra. Metanogenerna i C2, C4 och C6 är huvudsakligen hydrogenotrofa metanogener. Hydrolytiska fermentationsbakterier och acetogener bidrar båda till produktionen av ättiksyra och H2. De producerar också enzymer, kofaktorer och metaboliska signaler för att reglera väteproduktionen. Dessutom konkurrerar homoacetogena bakterier och acetogener inte i dessa områden. Hydrogenotrofa metanogener kan producera metan från CO2 och H2 som produceras i föregående steg. Därför används metaboliska vägar i dessa områden huvudsakligen för H2, formate och andra ämnen.
mer än 99% av C3-området har metylotrofa metanogener, såsom: Methanolobus. Brevibacter, Paenibacillus, Brochothrix och Lactococcus. Tidigare studier visade att metoxiaromatiska föreningar (en viktig del av lignocellulosa) bryts ned för att producera metanol och andra ämnen . Mikroorganismer i denna region kan bryta ner lignocellulosliknande materia av kol för att ge resurser till metylotrofa metanogener. Denna enkla mikrobiella gemenskap kan inte tillhandahålla tillräckliga substrat för metanogener som konsumerar H2. Biometanproduktionsvägen i detta område är baserad på konsumtionen av metylföreningar.
Staphylococcus detekterades också i C3-området. Nyligen isolerades Staphylococcus AntiMn-1 från djuphavssediment i Clarion-Clipperton-området med högt manganinnehåll. Den innehöll gener med hög resistens mot mangan, vilket tros vara en anpassning till den marina sedimentära miljön . Tungmetallhalten i C3-området är relativt hög. Det kan vara så att kolsömmiljön effektivt kan inducera uttryck av resistensgener, som kan ha antagonistiska och avgiftande effekter på transport och toxicitet hos tungmetaller inom mikroorganismer. Staphylococcus i detta område kan innehålla resistensgener för att anpassa sig till kolsömmiljön så att den också kan delta i fermentationsmetabolismen av kol. Det finns många olika arter av metanogener i C7, och de väteproducerande bakterierna domineras av Clostridium, Bacillus, Citrobacter och andra anaerober, som ger substrat för acetoklastiska metanogener och även H2, CO2 och formiat för hydrogenotrofa metanogener. Vidare reducerar ackumulerande ättiksyra sulfatreducerande bakterier, inklusive Desulfosporosinus och desulfitobacterium. SRB har en starkare affinitet för ättiksyra än acetoklastiska metanogener, men de konkurrerar inte med metylotrofa metanogener för vissa substrat, såsom metanol. Således kan metabolismen av både sulfatreducerande bakterier och metanogener fortsätta samtidigt i detta område . Metabolism i C7 dominerades av metylotrofa metanogener, följt av ättiksyrafermentering och sedan koldioxidreduktion.