Canal KcsA: Estructura
Este artículo se centrará en KcsA, un canal de potasio de Streptomyces lividans, que contiene dos dominios. La primera es la porción transmembrana, que contiene 120 residuos. Esto se analizará en detalle a continuación. El segundo dominio es un dominio citoplasmático que contiene alrededor de 40 residuos. El dominio citoplasmático contribuye a la estabilidad del estado cerrado. Esto se concluye en base a datos que muestran una disminución en la eficiencia del plegado y ensamblaje del canal y la estabilidad térmica que se ve afectada cuando el dominio citoplasmático se elimina de la estructura cristalina.5 Como se ve en la Figura 2, el dominio citoplasmático C-terminal lleno de agua es una estructura de haz de 4 hélices que se extiende linealmente hacia el citoplasma.5-6 Se ha demostrado por resonancia paramagnética electrónica, EPR, datos producidos por Uysal, que el C-terminal tiene un estiramiento que sobresale donde el dominio C-terminal se encuentra con el dominio transmembrana.5 Este tramo de escaleras que conecta los dos tiene un mayor grado de flexibilidad que las otras partes que potencialmente juega un papel durante la apertura del canal.5 Todavía hay mucha investigación que debe llevarse a cabo para comprender completamente la funcionalidad del dominio C-terminal. Una función sugerida del dominio es que puede servir como receptor para una proteína activadora plasmática no identificada o ligando, pero otros argumentan que no hay un efecto significativo en la ruta de permeación del ion. La estructura aún no contiene la orientación de ninguna cadena lateral, pero la disposición de las hélices sugiere interacciones electrostáticas en la subunidad.6 La Figura 3 muestra cómo se compara el dominio citoplasmático entre un canal abierto y un canal cerrado con dos canales bacterianos.7 El resto del artículo se centrará en el dominio transmembrana que selecciona el potasio.
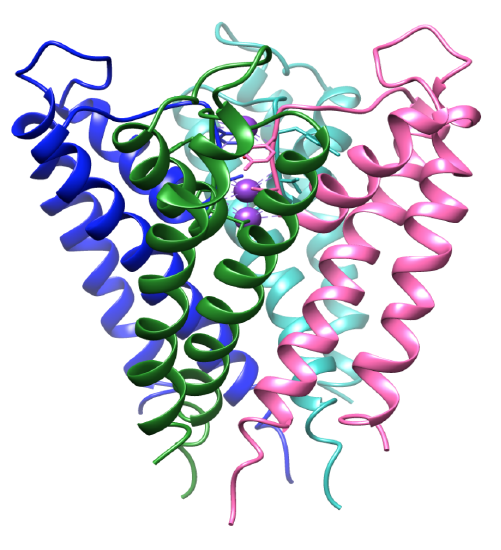
Figura 3. El dominio transmembrana. Cada subunidad tiene un color diferente y los iones de potasio son morados. AP 1BL8.
La selectividad para los iones de potasio depende en gran medida de la estructura del filtro de selectividad del canal de potasio. El entorno de coordinación del canal tiene ligandos selectivos que se unen específicamente a los iones de potasio.8 La proteína del canal de potasio es un tetrámero, que contiene cuatro subunidades idénticas que forman un poro central.8-9 Los iones fluyen hacia un poro que se abre y se cierra, dirigiendo los iones a la cavidad central. Las subunidades contienen dos hélices alfa donde una se enfrenta al poro central, la hélice interna, y la otra se enfrenta a la membrana lipídica, también conocida como la hélice externa.3 Junto con las hélices interior y exterior, cada subunidad incluye una hélice de poros.3,8 Las subunidades están inclinadas, como se ve en la Figura 3, donde el filtro de selectividad está cerca de la superficie extracelular de la membrana. Las cuatro hélices internas están estructuradas de una manera en la que se unen cerca de la superficie intracelular. Tanto las entradas intra como extracelulares están cargadas negativamente por aminoácidos ácidos. Esto permite una mayor concentración de cationes cerca de la membrana y menos aniones debido a las cargas opuestas.8

Figura 4. Canal de potasio (estructura KcsA de Streptomyces lividans) con componentes estructurales clave etiquetados. Los principales aspectos a tener en cuenta son el filtro de poros, cavidad y selectividad (SF). Estas son las partes del canal por donde viajan los iones de potasio. Primero, el ion fluye a través del poro y entra en la cavidad hidratado cuando el poro está abierto. Luego se deshidrata un ion de potasio y se selecciona para viajar a través del SF, donde luego ingresará a la célula. Tenga en cuenta que solo se muestran dos subunidades en esta imagen para facilitar la visualización. PBD 1K4C.
La región de los poros se identificó por primera vez con toxinas de escorpión que bloquean los poros. Interactúan con los aminoácidos en la entrada del poro causando bloqueo y disfunción. El poro es donde entra el ion de potasio, donde luego se transfiere a la cavidad central, como se ve en la Figura 4. Durante este tiempo, el ion de potasio permanece hidratado.8 El poro intracelular es la característica del canal que permite que los iones de potasio entren en la célula cuando está abierta, o que se bloquee la entrada cuando está cerrada. Los canales se abren cuando hay un cambio en el potencial de membrana, lo que permite la entrada de iones de potasio en el filtro de cavidad. Una vez que se produce la unión selectiva en el filtro de selectividad, el potencial cae a través de la membrana, cerrando así el poro.1 El poro y la cavidad están revestidos hidrofóbicamente, lo que permite reducir la barrera electrostática. Debe haber una barrera electrostática más baja para que el costo energético de la deshidratación se compense debido a la gran barrera de energía para la permeación iónica (barrera dieléctrica). Por lo tanto, la cavidad llena de agua permite un entorno energéticamente favorable.3,8,10,11
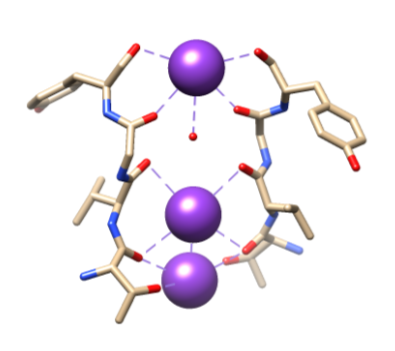
Figura 5. Unión de iones de potasio (púrpura) a ligandos específicos por las O troncales de Thr 75, Val 76, Gly 77 y Tyr 78. La coordinación completa no se muestra aquí para facilitar la visualización. Reproducido de Samsom.11
A continuación, los iones deben seleccionarse en el filtro de selectividad (SF), que está revestido con átomos de cadena principal polar que pertenecen a aminoácidos. El FS es tan estrecho que los iones deben deshidratarse antes de entrar en él. Hay dos características esenciales en el SF. En primer lugar, los átomos de la cadena principal crean dimensiones adecuadas para coordinar un ion de potasio deshidratado mediante apilamiento, formando así anillos de oxígeno secuenciales. Esto crea una distancia muy pequeña que el ion de potasio debe recorrer para llegar a cada sitio mientras viaja a través del filtro. Esto forma una secuencia de cuatro sitios de coordinación óctuplos cúbicos, como se ve en la Figura 6.3,8 En cualquier momento, estos sitios están ocupados por dos iones K+ y dos moléculas de agua en una secuencia alterna La segunda característica es el empaquetamiento de proteínas alrededor del SF, que sigue una secuencia de T-V-G-Y-G, mostrada en la Figura 5.9 La conducción puede ocurrir cuando dos iones de potasio entran en el SF, causando un equilibrio entre la fuerza de atracción que ocurre desde el SF al ion con la fuerza de repulsión que ocurre entre los dos iones de potasio.8 Esta repulsión ayuda a los iones de potasio a superar su afinidad intrínseca para unirse firmemente a los sitios de unión.3 A continuación se analizará más a fondo el mecanismo de selección.
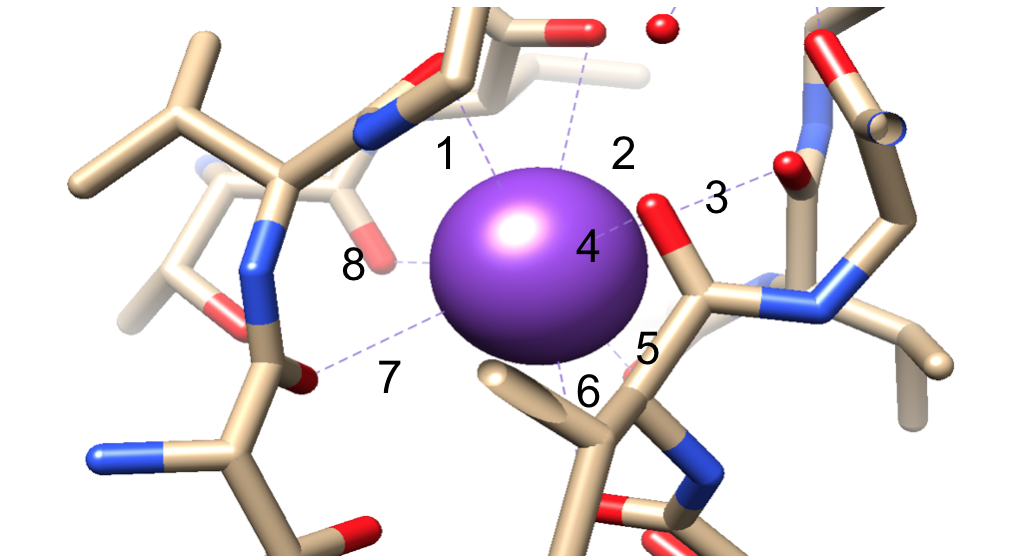
Figura 6. Un ejemplo de un ion de potasio (púrpura) en un sitio de coordinación óctuple. Encuadernación numerada para facilitar la visualización.