vita
itt 25 chironomid faj genomméretét határoztuk meg; ezek az értékek egybevágtak a szakirodalomban korábban közölt három faj genomméretével (Petitpierre, 1996; Schmidt-Ott et al., 2009), amelyek 0,13 és 0,21 pg DNS között voltak. A tanulmány eredményei azt mutatták, hogy a chironomidák genommérete egyenletesen nagyon kicsi volt (ábra. 3, 1. táblázat). A Chironomidae családra kapott átlagos C-érték 0,13 + 0.03 pg DNS, amely egyértelműen kisebb, mint a D. melanogaster genom mérete. A rovarok közül a legkisebb genomméretet a Strepsiptera Caenocholax fenyesi körülbelül 0,11 pg DNS-sel (ami nagyjából 108 Mb-nak felel meg), bár a hesseni légy Mayetiola destruktor azt is javasolták, hogy még kisebb genommérete 0,09 pg legyen (Johnston et al., 2004; Gregory, 2014). Itt legalább három fajról számolunk be, amelyek genomja kisebb, mint bármely ismert rovaré: C. tsushimensis, Diamesa japonica és Hydrobaenus tsukubalatus, C-értéke 0,07 pg, 0,08 pg és 0,08 pg (1.táblázat). Tudomásunk szerint C. tsushimensis (ábra. 1B) ma már úgy tekinthető, hogy az ismert rovarok közül a legkisebb genommérettel rendelkezik, C-értéke 0,07 pg DNS, ami nagyjából 68 Mb-nak felel meg. Ez az érték egybevág egy rokon faj, a Clunio marinus genomméretével, amelyet egyszer 95 Mb-ra, újabban 87,2 Mb-ra becsültek (Kaiser and Heckel, 2012; Tobias S. Kaiser, personal communication).
a közelmúltban az antarktiszi midge, B. Antarktisz genomját szekvenálták (Kelley et al., 2014). A szerzők azt állították, hogy ennek a fajnak a genommérete 89-re becsülhető.5-105 Mb volt a legkisebb az összes rovar között. Itt azonban megmutatjuk, hogy ez a genomméret a chironomidák normál tartományán belül van értékek. A szerzők azt is felvetették, hogy a B. Antarktisz kis genommérete alkalmazkodott a szélsőséges hideg környezethez. Adataink megerősíthetik ezt a hipotézist, mivel a D. japonica, a Diamesinae alcsaládból származó alpesi faj is apró genomméretet mutat. Egy másik kapcsolódó Diamesinae, a Himalája midge Diamesa sp. valójában -16 C-os hőmérsékleten aktívnak bizonyult (Kohshima, 1984). Azonban a telmatogeton japonicus, amely nagyon alacsony hőmérsékleten is képes fejlődni (Danks, 1971; Sunose és Fujisawa, 1982), viszonylag nagy genomméretet mutat a chironomid család átlagához képest. Itt figyelembe kell venni a genomméretre gyakorolt filogenetikai hatást, mivel a B. Antarktisz az Orthocladiinae alcsaládba tartozik, amely átlagosan kis genomméretet mutat. A C. tsushimensis és a Hydrobaenus tsukubalatus fajok, amelyek a legkisebb genomméretet mutatják a chironomidák között, szintén az Orthocladiinae alcsaládba tartoznak. Így a B kis genommérete. az Antarktisz egyszerűen egy ősi tulajdonság, nem pedig a szélsőséges hideg alkalmazkodás eredménye.
a P. vanderplanki az egyetlen rovarfaj, amelyről ismert, hogy teljesen dehidratált állapotban éri el az anhidrobiózist (ábra. 1D) és regenerálódik a rehidráció után (Hinton, 1951, 1960A; Watanabe, 2006; Cornette and Kikawada, 2011). Kutatócsoportunk nemrég publikálta a kiszáradástűrő Polypedilum vanderplanki és a kiszáradásra érzékeny congeneric Polypedilum nubifer genomszekvenciáinak összehasonlító elemzését (Gusev et al., 2014). Az összehasonlítás azt mutatta, hogy mindkét faj genommérete hasonló volt, és csak korlátozott számú duplikált géncsoport kapcsolódott a P. vanderplanki anhidrobiózisához (Gusev et al., 2014). A genom összeállításából levezetett genomméretek a P. vanderplanki esetében 104 Mb, a P. nubifer esetében pedig 107 Mb voltak, és ezek az értékek nem különböztek jelentősen a jelen vizsgálatban e fajok esetében mért genomméretektől, ami körülbelül 98 Mb-nak felelt meg. P. vanderplanki és P. a nubifer genomok hasonló alacsony DNS-ismétlődési arányt mutattak, és csak kis számú átültethető elemet találtak, összehasonlítva más dipterán fajokkal. Az átültethető elemek arányának ez a csökkenése az antarktiszi midge-ben is megfigyelhető volt, B. Antarktisz (Kelley et al., 2014). Az átlagos intronhossz jelentősen csökkent az antarktiszi törpében, valamint mind P. vanderplanki, mind P. nubifer esetében (Gusev et al., 2014). Mivel a kiszáradás érzékeny és fagyasztva intoleráns P. a nubifer osztja ezeket a tulajdonságokat az antarktiszi törpével és az anhidrobiotikus törpével, a transzponálható elemek alacsony aránya és a rövid intronhossz az összes chironomidának közös jellemzője lehet, és lehet, hogy nem kapcsolódik közvetlenül a szélsőséges környezetekkel szembeni toleranciához, még akkor sem, ha az adaptív hatás nem zárható ki.
ábra. 3.
a Diptera törzsfejlődése becsült C-értékeket mutat. A Chironomid család fekete színnel látható. A kötőjel vonal a D. melanogaster genomméretét mutatja. A filogenetikai kladogram különböző Chironomid és Dipteran fák konszenzusa (Yeates and Wiegmann, 1999, 2005; Saether, 2000; Cranston et al., 2012). A hibák sávjai szórást mutatnak. A hibasávok nélküli genomméreteket az állati Genomméret-adatbázisban felsorolt korábbi vizsgálatokból nyertük (Gregory, 2014).
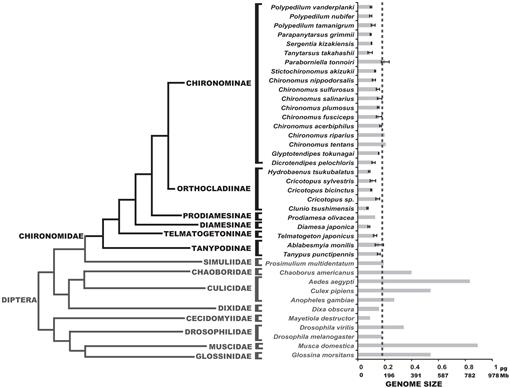
Fig. 4.
a testméret és a genomméret közötti kapcsolat a jelen vizsgálatban vizsgált chironomid Fajok között. A) minden faj esetében az átlagos teljes testhossz milliméterben, az átlagos C-érték pedig pg DNS-ben van kifejezve. A két változó között csak gyenge pozitív korrelációt (r2 = 0,171, n = 25) figyeltek meg, amelyet szilárd vonal valósít meg. (B) a nemzetség szintjén az azonos értékű eszközöket filogenetikailag független kontrasztok módszerével alakítottuk át, Cranston et al. (2012). A transzformált kontrasztok közötti korreláció, amelyet egy folytonos vonal valósít meg, nem volt szignifikáns (r2 = 0,118, n = 13). Az adatelemzést a Prism 4-en végezték.0 szoftver Macintosh – hoz (GraphPad Software, Inc.).

milyen adaptív tulajdonságok lehetnek potenciálisan társítva a Chironomidae család apró genomméreteivel? Valójában számos fejlődési és ökológiai tényezőt javasoltak, hogy korreláljanak a genom méretével. Először is, számos taxonban találtak pozitív korrelációt a genomméret és a testméret között, és ez a korreláció viszonylag általános jelenségnek tűnik (Gregory, 2005). Ezt az összefüggést a szúnyogoknál is jelentették (Ferrari and Rai, 1989), de más rovar taxonoknál nem mindig egyértelmű (Gregory and Hebert, 2003; Ardila-Garcia and Gregory, 2009). A chironomidákon belül csak gyenge pozitív korrelációt találtunk a genom mérete és a testméret között (ábra. 4A). A Chironomus nemzetség átlagos C-értéke (0,15 pg) magasabb volt, mint a Polypedilum nemzetségé (0,10 pg), és ez a különbség a Chironomus fajok általában nagyobb testméretének tudható be. A két nemzetség közötti filogenetikai távolság azonban megmagyarázhatja ezt a genomméret-különbséget is. Következésképpen a nemzetségi szintű adatainkat filogenetikailag független kontrasztelemzéssel korrigáltuk (Garland and Adolph, 1994; Garland et al., 2005), és ennek eredményeként a genomméret és a testméret közötti összefüggés nem szignifikánsnak bizonyult (ábra. 4B). A Chironomidák genomméretének szűk tartománya (csak két—háromszoros) és ezeknek a fajoknak az átlagos kis testmérete (a legtöbb nem haladja meg a néhány millimétert) valószínűleg megnehezíti a Chironomidae családon belüli mindkét tulajdonság közötti összefüggés megtalálását.
a rovarok között egy másik összefüggést javasoltak a genom mérete és a fejlődési komplexitás között. Míg a hemimetabolous rovarok, csak fokozatos nymphal moltokkal, a genomméretek széles skáláját mutatják (C-értékek 0,18-tól 16-ig.93 pg), a komplex metamorfózisukkal rendelkező holometabolos rovarok C-értékei a feltételezett 2 pg küszöbön belül korlátozottak, többségük esetében (Gregory, 2002, 2005). A fejlődési komplexitás hatása a genom méretére itt nyilvánvaló, mivel a holometabolos rovarok intenzív morfológiai átalakításon mennek keresztül a metamorfózis korlátozott ideje alatt, a kis genomméret pedig előnyöket jelent a sejtosztódás nagy sebességű megvalósításához. A chironomidák holometabolos rovarok, metamorfózisuk rendkívül gyors. Pupilla stádiumuk csak egy napig tarthat, vagy bizonyos fajok esetében akár csak néhány órát is igénybe vehet (Cranston, 1995c). Például a C. tsushimensis komplex metamorfózist mutat kifejezett szexuális dimorfizmussal (ábra. 1B).
a genomméret és a sejtosztódás sebessége közötti kapcsolat szintén befolyásolja a fejlődési sebességet. Például azt javasolták, hogy a levéltetvek gyors életciklusa összefüggésbe hozható kis genomméretükkel (Ma et al., 1992). Így a kis genomméreteknek előnyt kell jelenteniük az ideiglenes környezetben gyorsan fejlődő chironomid fajok számára. A Clunio nemzetségbe tartozó chironomidák átmeneti dagálymedencékké fejlődhetnek, és a felnőttek szinkronizált megjelenését is mutatják a holdciklushoz viszonyítva és az apály rövid ideje alatt (Kaiser and Heckel, 2012). Ennek eléréséhez magas fejlődési arányra van szükség. Vegye figyelembe, hogy a hasonló intersticiális környezetben gyorsan fejlődő kis polychaete annelidek kisebb genomméreteket mutatnak, mint a makrobentikus Fajok (Gambi et al., 1997). Antarktiszi szúnyogok, például B. Antarktisz vagy eretmoptera murphyi (Lee et al., 2006; Worland, 2010), a lárva fejlődése két évig tart, többnyire lefagyott állapotban, de a metamorfózisnak és a szaporodásnak a nagyon rövid nyári időszakban kell bekövetkeznie. Ebben az esetben magas fejlődési sebességre van szükség, amelyet elősegíthet a B. Antarktisz kis genomja. Ezt a jelenséget az angiospermium növényekről szóló tanulmány szemlélteti, amely azt mutatja, hogy azok a fajok, amelyek képesek teljes életciklusukat befejezni a rövid Antarktiszi nyár alatt, bemutatták a legkisebb genomméreteket (Bennett et al., 1982). Egy másik szélsőség, az Anhidrobiotikus midge P. vanderplanki az efemer kőzetmedencékhez igazodik. Itt is, az ideiglenes élőhely magas fejlődési üteme összekapcsolható e faj kis genomméretével. Az Ausztrál faj azonban P. tonnoiri, amely szintén kiszáradásnak van kitéve hasonló sziklamedencéken (Jones, 1975; Adams, 1985; Frouz et al., 2003), viszonylag nagy genomméretet mutat egy chironomid számára (ábra. 3. és 1. táblázat).
az abiotikus stressz valószínűleg befolyásolja a DNS integritását és ennek következtében a genom méretét is. Például a P. lárvái. a vanderpianki az anhidrobiózis után hatalmas DNS-károsodást tapasztal, és a hatékony DNS-javítás a rehidrációt követő néhány nap alatt történik (Gusev et al., 2010). A növényekben negatív korrelációt találtak a genom mérete és a sugárzási tolerancia között (Bennett and Leitch, 2005), és a mutációs ráta alacsonyabb volt a kisebb genomméretű fajoknál (Abrahamson et al., 1973). Következésképpen a P. vanderpianki kis genommérete előnyt jelenthet a káros mutációk felhalmozódásának elkerülésére a kiszáradási és rehidrációs ciklusokhoz kapcsolódó DNS-javítási események során. Tengeri fajként a C. tsushimensis magas sótartalmú stressznek is ki van téve, amely a legtöbb rovarfaj számára halálos. A magas sótartalom és más abiotikus stresszek ismert módon intracelluláris reaktív oxigénfajokat (ros) generálnak, amelyeket a DNS károsodásának fő forrásaként javasoltak (Franca et al., 2007; Gill és Tuteja, 2010). Ez összefügghet a kis genommérettel is. Más tengeri és magas sótartalmú fajok, mint például a T. japonicus vagy a Chironomus salinarius nem mutatnak olyan kicsi genomméretet, mint a C. tsushimensis. Savtűrő fajok, például Chironomus sulfurosus, Chironomus acerbiphilus vagy Polypedilum tamanigrum (ábra. 1C) képes túlélni olyan forró forrásokban, amelyek pH-ja olyan alacsony, mint 1,4 (Doi et al., 2004; Takagi et al., 2005). Itt is nem volt egyértelmű összefüggés a savas stresszel szembeni tolerancia és a kis genomméret között, és a filogenetikai hatás fontosabbnak tűnt, mivel a Polypedilum nemzetség kisebb genommérete és a Chironomus nemzetség nagyobb genommérete volt.
összefoglalva, a kironomidákban megfigyelt apró genomméretek valószínűleg az átültethető elemek számának és a genomszerkezetükben lévő intronok hosszának ősi csökkenésének következményei. A Chironomidae családban a genomméretek a törzsfejlődés szerint különböztek az alcsalád és a nemzetség szintjén. A különböző taxonok összefüggéseiből következtetve, a kironomidok kis genommérete elméletileg az instabil és szélsőséges környezetekhez való alkalmazkodást jelenthette a magas fejlődési ráta és az alacsony mutációs ráta miatt. Reméljük, hogy ez a tanulmány ösztönözni fogja a Chironomid Genom jövőbeli projektjeit, és hogy az összehasonlító genomika ezen a taxonon belül segít megérteni, hogy a Chironomid genomok hogyan lépnek kölcsönhatásba a környezetükkel.