1. Eigenschaften
2. Funktionen
– Turgor
– Lagerung
– Lytisch
– Apoptose
– Andere
3. Vesikulärer Transport
Vakuolen sind membrangebundene Organellen, die in Pflanzenzellen und Pilzen, einschließlich Hefen, vorkommen. Sie sind kritische Organellen für die Funktion von Pflanzenzellen.
Merkmale
Vakuolen sind normalerweise große Kompartimente, die in reifen Zellen bis zu 90 % des gesamten Zellvolumens ausmachen können (Abbildungen 1 und 2). Sie sind das größte Kompartiment von Pflanzenzellen. Der Name Vakuole leitet sich vom lateinischen Wort „vacuus“ ab, was leer bedeutet. Dies war eindeutig ein Missverständnis, da Vakuolen nicht leer sind, sondern mit einer mehr oder weniger konzentrierten wässrigen Lösung gefüllt sind. Die Membran der Vakuole ist als Tonoplast bekannt und ein wesentlicher Bestandteil für die Funktion dieser Organelle. In Pflanzen gibt es verschiedene Arten von Vakuolen, je nachdem, welche Rolle sie spielen. Eine Pflanzenzelle kann verschiedene Arten von Vakuolen enthalten, und eine Vakuole kann ihr Enzymrepertorium modifizieren und dann ihre Funktion ändern.
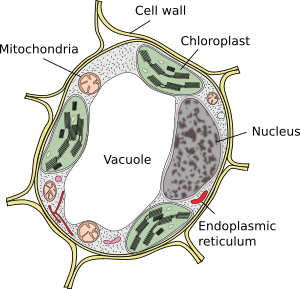
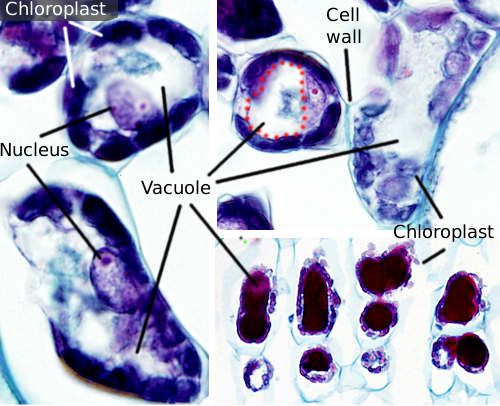
Vakuolen sind normalerweise abgerundet, aber die endgültige Form wird durch die Zellmorphologie beeinflusst. Eine große Vakuole wird oft in reifen Pflanzenzellen beobachtet. Die Membran der Vakuole wird jedoch manchmal tief und stark gefaltet und bildet kleine Kompartimente, die bei Lichtmikroskopie wie viele kleine Vakuolen aussehen, aber tatsächlich nur eine Vakuole sind, da die Membran kontinuierlich ist.
Neue Vakuolen werden durch Fusion von Vesikeln gebildet, die aus dem Golgi-Apparat freigesetzt werden. Zunächst bilden sie ein neues Kompartiment, das als Pro-Vakuole bekannt ist. Eine meristematische Zelle kann Hunderte von Provakuolen haben. Während der Zelldifferenzierung verschmelzen dann Pro-Vakuolen zu kleinen Vakuolen, und der Fusionsprozess wird fortgesetzt, bis eine große zentrale Vakuole gebildet wird. Das endoplasmatische Retikulum könnte auch an der Bildung und dem Wachstum von Vakuolen in einigen Pflanzenzellen, meist in Samen, beteiligt sein. Sobald eine große Vakuole vorhanden ist, regulieren Vesikel aus dem Golgi-Apparat und der Plasmamembran die Größe durch Hinzufügen und Entfernen der Membran.
Die Hauptvakuole der meisten Pflanzenzellen ist ein großes Kompartiment, das mit einer sauren Lösung gefüllt ist, die Salze (Natrium, Kalium), Metaboliten (Kohlenhydrate, organische Säuren) und einige Pigmente enthält. Einige dieser Moleküle gelangen aus dem Cytosol gegen Konzentrationsgradienten in die Vakuole. Der normale pH-Wert in der Vakuole liegt zwischen 5 und 5, 5, obwohl er in der Zitronenfrucht bei etwa 2 oder in einigen Algen sogar bei 0, 6 liegen kann.
Funktion
Vakuolen sind essentiell für die Physiologie und Homöostase pflanzlicher Zellen und erfüllen je nach Zelltyp unterschiedliche Funktionen. Die folgenden sind einige von ihnen:
Turgor
Zellturgor ist das Niveau des hydrostatischen Drucks gegen die Zellwand der Pflanzenzelle. Dieser Druck steht unter der Kontrolle von Vakuolen, die verschiedene Substanzen, einschließlich Ionen, in sich aufnehmen, um im Vergleich zu denen des Cytosols variable innere osmotische Umgebungen zu erzeugen. Durch die unterschiedliche Osmolarität auf beiden Seiten der Vakuolenmembran durchquert das Wasser die Membran entweder nach innen oder nach außen. Die Substanzen, die zur Vakuolenosmolarität beitragen, können die Vakuolenmembran durch ATP-abhängigen Transport durchqueren, der durch Ionenpumpen vermittelt wird. H (+) -ATPase und H (+) -Pyrophosphatase können Protonengradienten zwischen beiden Seiten der Vakuolenmembran bilden, und diese Gradienten werden zum Transport anderer Moleküle verwendet. Die Fähigkeit, Wasser in der Vakuole zu speichern, ist für das Wachstum von Pflanzenzellen nach der Mitose unerlässlich. Pflanzenzellen können ihre Größe um das 10- bis 20-fache erhöhen, was für das Wachstum der Körperpflanze und für die Veränderung der Form von Pflanzenorganen sehr nützlich ist. Das durch hydrostatischen Druck vermittelte Wachstum spart Energie, da es billiger ist, die Wassermenge zu erhöhen, als neue Moleküle zu synthetisieren (tierisches Zellwachstum basiert auf molekularer Synthese). Für Pflanzenzellen ist es sicherer, Wasser in der Vakuole anzusammeln, da auf diese Weise die cytosolischen Moleküle nicht verdünnt werden, was das Überleben der Zellen beeinträchtigen würde.
Lagerung
Vakuolen sind die letzte Station für einige vesikuläre Verkehrswege. In einigen Zellen speichern sie Kohlenhydrate und Proteine. Dies geschieht eindeutig in Samen, wo Vakuolen Proteine ansammeln, die während der Keimung benötigt werden. Speichervakuolen werden während der Zelldifferenzierung zu lytischen Vakuolen. Im Gegensatz zu Tieren haben Pflanzen kein Ausscheidungssystem und können sich auch nicht bewegen, um giftige Substanzen zu vermeiden. In Pflanzen werden potenziell gefährliche Substanzen in Vakuolen gelagert. Auf diese Weise werden Stoffwechselrückstände und toxische Substanzen wie Schwermetalle (Cadmium, Zink und Nickel) in Vakuolen gefunden. Darüber hinaus speichern sie auch andere Substanzen wie Pigmente (z. B. Anthocyane) in den Epidermiszellen von Blütenblättern, toxische Substanzen gegen Pflanzenfresser, Harze, Alkaloide wie Opium usw. Der größte Teil des Geschmacks von Obst und Gemüse ist das Ergebnis von Substanzen, die in Vakuolen gelagert werden.
Abbauzentren
Lytische Vakuolen können in vegetativen Geweben gefunden werden, daher werden sie auch als vegetative Vakuolen bezeichnet. Sie enthalten Enzyme wie Proteasen und Nukleasen sowie eine Reihe von Proteinen, die an der Abwehr von Krankheitserregern beteiligt sind. Protonenpumpen, die in die Vakuolenmembran eingesetzt sind, geben Protonen in die Vakuole ein und säuern den Inneninhalt an. Der niedrige pH-Wert und die lytischen Enzyme ermöglichen Abbauprozesse. Vakuolen spielen eine ähnliche Rolle wie Lysosomen tierischer Zellen. Darüber hinaus sind Vakuolen wie Lysosomen an der Autophagie beteiligt. Vakuolare Verarbeitungsenzyme sind Proteine, die auch in Vakuolen vorkommen. Sie wandeln molekulare Vorläufer, die als inaktive Moleküle in die Vakuole gelangen, in aktive Moleküle um.
Apoptose
Vakuolen sind über einen Mechanismus, der als Autolyse bekannt ist, an der Apoptose von Pflanzenzellen beteiligt. Darüber hinaus tritt eine Art von Zelltod, der als überempfindlicher Zelltod bekannt ist, in Pflanzenzellen auf, wenn die Vakuolenmembran gebrochen wird.
Andere
Es gibt spezialisierte Vakuolen in verschiedenen Pflanzengeweben. Zum Beispiel akkumulieren Vakuolen in den inneren Tegumenten des Samens Flavonoide zum Schutz vor ultraviolettem Licht. Flavonoide werden in der cytosolischen Oberfläche der endoplasmatischen Retikulummembranen synthetisiert und dann zur endgültigen chemischen Verarbeitung in das Innere der Vakuolen transloziert. In der Vakuolenmembran befinden sich Transporter, um diese Translokation durchzuführen.
Einige Pflanzenarten, wie Brassicas, haben Vakuolen in ihrem vegetativen Gewebe, um Pflanzenfresser aufzuhalten. Diese Vakuolen speichern Proteine wie Myrosinasen. Sobald diese Enzyme durch die Pflanzenfresseraktivität freigesetzt werden, bauen sie molekulare Verbindungen der Blätter ab, die für das Tier giftig werden. Zellen, die Myrosin speichern, sind als Myrosinzellen bekannt und befinden sich in der Nähe der Gefäßbündel der Blätter.
Pflanzen haben kein Immunsystem, so dass jede Zelle ihr eigenes Abwehrsystem hat. Abwehrproteine und Enzyme können in Vakuolen gefunden werden. Es gibt zwei Abwehrmechanismen, die Vakuolen ausführen können (Abbildung 3): Vakuolenmembran kollabiert und Fusion der Membrankolonne mit der Plasmamembran. Virusinfektionen führen zum Bruch der Vakuolenmembran und setzen Enzyme in das Cytosol frei, wo sie Viren angreifen können. Die Fusion von Vakuolenmembran und Plasmamembran setzt Vakuolenenzyme in den extrazellulären Raum frei, wo sie Krankheitserreger wie Bakterien abtöten können.
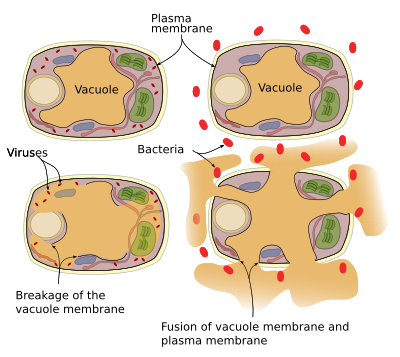
Vesikulärer Verkehr
Vakuolen sind Teil des vesikulären Verkehrs. Tatsächlich können sie als Endprodukt des vesikulären Transports angesehen werden, da ihre Bildung und Aufrechterhaltung von den ankommenden Vesikeln abhängt. Moleküle, die gespeichert oder abgebaut werden sollen, einschließlich hydrolytischer Enzyme, sowie alle Membranmoleküle werden über Vesikel auf Vakuolen gerichtet. Moleküle können verschiedenen vesikulären Wegen folgen, um zu Vakuolen zu gelangen:
Endoplasmatisches Retikulum> Golgi-Apparat > Vakuole; Golgi-Apparat> prävakuolares Kompartiment > Vakuole. Dies ist der Standardweg, um hydrolytische Enzyme in Richtung Vakuolen zu transportieren. Prävakuoläre Kompartimente ähneln multivesikulären Körpern / späten Endosomen tierischer Zellen. Seltsamerweise werden hydrolytische Enzyme im Golgi-Apparat nicht durch 6-Phosphat-Mannose-Einheiten ausgewählt, sondern durch eine Sequenz von Aminosäuren, die sich in ihrer Aminosäurekette befinden. Es gibt spezifische Sequenzen von Aminosäuren, um Proteine auf die hydrolytischen Vakuolen abzuzielen, und andere Sequenzen sind spezifisch, um andere Proteine in Richtung der Speichervakuolen zu bewegen. Alle Proteine, die auf Vakuolen abzielen, haben eine Signalsequenz und müssen von Rezeptoren spezifisch erkannt werden.
Endoplasmatisches Retikulum> Vakuole. Moleküle können direkt aus dem endoplasmatischen Retikulum in Vakuolen gelangen. Dieser Weg ist in Samen als Weg zur Lagerung prominent. In anderen Pflanzenzellen, wie in Blättern, kann dieser Weg jedoch sehr selten sein. Vesikel, die vom endoplasmatischen Retikulum zu Vakuolen wandern, sind unabhängig von COP-II-Schichten, die für Vesikel benötigt werden, die auf den Golgi-Apparat abzielen. Im endoplasmatischen Retikulum-Vakuolen-Weg gibt es manchmal intermediäre Kompartimente, aber sie sind transiente membrangebundene Organellen, in denen Moleküle kurz zurückgehalten werden, bevor sie in die Vakuole gelangen. Dieser vesikuläre Weg kann von zellulären Autophagie-Komponenten abgeleitet sein.
Plasmamembran > Vakuole. Endozytäre Vesikel verschmelzen direkt mit Vakuolen, die wie frühe Endosomen funktionieren.
Bibliographie
Marty F. 1999. Pflanzenvakuolen. Pflanzenzelle 11:587-600.
Pereira C., Pereira S, Pissarra J. 2014. Abgabe von Proteinen an die Pflanzenvakuole – ein Update. Internationale Zeitschrift für molekulare Wissenschaften 15: 7611-762.
Shimada T, Takagi J, Ichino T, Shirakawa M, Hara-Nishimura I. 2018. Pflanzenvakuole. In: Annual review in plant biology. https://doi.org/10.1146/annurev-arplant-042817-040508.
Taiz L. 1992. Die Pflanzenvakuole. Zeitschrift für experimentelle Biologie 172: 113-122.
Zhang C, Hicks G R, Raikhel NV. 2014. Pflanzenvakuolenmorphologie und Vakuolentransport. Grenzen in den Pflanzenwissenschaften 5: 476. ![]()