1. Jellemzők
2. Funkciók
– Turgor
– Tárolás
– Litikus
– Apoptózis
– Egyéb
3. Hólyagos kereskedelem
a Vakuolok membránhoz kötött organellák, amelyek növényi sejtekben és gombákban találhatók, beleértve az élesztőket is. Ezek kritikus organellák a növényi sejtek működéséhez.
jellemzők
a Vakuolok általában nagy rekeszek, amelyek érett sejtekben a teljes sejttérfogat 90% – át teszik ki (1.és 2. ábra). Ezek a növényi sejtek legnagyobb rekesze. A vacuole név a Latin “vacuus” szóból származik, ami üres. Ez egyértelműen félreértés volt, mert a vakuolok nem üresek, hanem többé-kevésbé koncentrált vizes oldattal vannak feltöltve. A vakuol membránja tonoplaszt néven ismert, és elengedhetetlen része ennek az organellának a működéséhez. A növényekben többféle vakuol létezik az általuk betöltött szerep szerint. Egy növényi sejt különböző típusú vacuolákat tartalmazhat, a vacuol pedig módosíthatja enzim repertoárját, majd megváltoztathatja funkcióját.
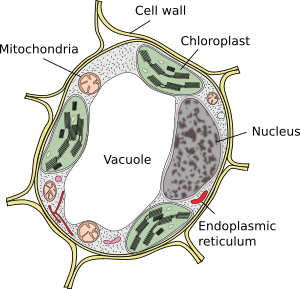
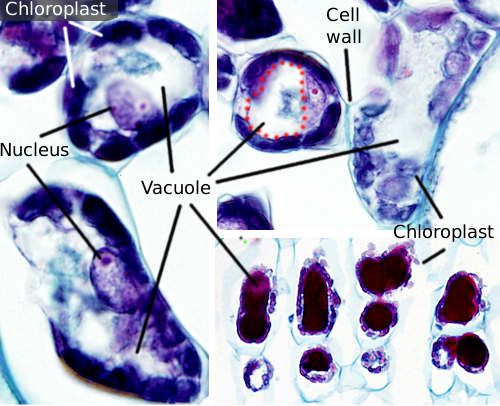
a Vakuolák általában lekerekítettek, de a végső alakot a sejtmorfológia befolyásolja. Egy nagy vakuolát gyakran megfigyelnek az érett növényi sejtekben. Azonban a vakuol membránja néha mélyen és bőségesen összehajtódik, és kis rekeszeket képez, amelyek sok kis vakuolának tűnnek, amikor fénymikroszkóppal figyelik meg, de valójában csak egy vakuolák, mert a membrán folyamatos.
új vakuolák képződnek a Golgi-készülékből felszabaduló vezikulák fúziójával. Kezdetben egy új rekeszt alkotnak, amelyet pro-vacuole néven ismernek. Egy merisztematikus sejtnek több száz pro-vacuolája lehet. Ezután a sejtdifferenciálás során a pro-vacuolák kis vacuolákká olvadnak össze, és a fúziós folyamat addig folytatódik, amíg egy nagy központi vacuol képződik. Az endoplazmatikus retikulum részt vehet a vacuolák kialakulásában és növekedésében egyes növényi sejtekben, főleg a magokban. Ha egy nagy vakuol jelen van, a Golgi-készülékből és a plazmamembránból származó vezikulák szabályozzák a méretet a membrán hozzáadásával és eltávolításával.
a legtöbb növényi sejt fő vakuolája egy nagy rekesz, amely sókat (nátrium, kálium), metabolitokat (szénhidrátok, szerves savak) és néhány pigmentet tartalmazó savas oldattal van feltöltve. Ezen molekulák egy része a citoszolból jut be a vakuolába a koncentrációgradiens ellen. A vakuolán belüli normál pH 5 és 5,5 között mozog, bár a citromgyümölcsben 2 körül lehet, vagy egyes algákban akár 0,6 is lehet.
funkció
a Vakuolok elengedhetetlenek a növényi sejtek fiziológiájához és homeosztázisához, és a sejttípusnak megfelelően különböző funkciókat látnak el. A következők közül néhány:
Turgor
a sejt turgor a növényi sejt sejtfalához viszonyított hidrosztatikus nyomás szintje. Ez a nyomás a vakuolák ellenőrzése alatt áll, amelyek különböző anyagokat juttatnak be, beleértve az ionokat is, hogy változó belső ozmotikus környezetet hozzanak létre, összehasonlítva a citoszoléval. A vakuolamembrán mindkét oldalán eltérő ozmolaritás miatt a víz befelé vagy kifelé halad a membránon. Azok az anyagok, amelyek hozzájárulnak a vakuol ozmolaritásához, az ionos szivattyúk által közvetített ATP-függő transzport révén átjuthatnak a vakuol membránon. A H (+)-ATPáz és a H (+) – pirofoszfatáz képes protongradienseket képezni a vakuolamembrán mindkét oldala között, és ezeket a gradienseket más molekulák szállítására használják. A víz tárolásának képessége a vakuolában elengedhetetlen a növényi sejtek mitózis utáni növekedéséhez. A növényi sejtek 10-20-szorosára növelhetik méretüket, ami nagyon hasznos a testnövény növekedéséhez és a növényi szervek alakjának módosításához. A hidrosztatikus nyomás által közvetített növekedés energiát takarít meg, mivel olcsóbb a víz mennyiségének növelése, mint az új molekulák szintetizálása (az állati sejtek növekedése molekuláris szintézisen alapul). A növényi sejtek számára biztonságosabb a víz felhalmozódása a vakuolában, mert ily módon a citoszolos molekulák nem hígulnak fel, ami veszélyeztetné a sejtek túlélését.
Tárolás
a vakuolok néhány vezikuláris forgalmi útvonal utolsó állomása. Egyes sejtekben ezek a rekeszek a szénhidrátok és fehérjék tárolására. Ez egyértelműen a magokban történik, ahol a vacuolák felhalmozódnak a csírázás során szükséges fehérjékben. A tárolási vakuolok litikus vakuolákká válnak a sejtek differenciálódása során. Az állatoktól eltérően a növények nem rendelkeznek ürülékrendszerrel, és nem tudnak mozogni a mérgező anyagok elkerülése érdekében. A növényekben a potenciálisan veszélyes anyagokat vacuolákban tárolják. Ily módon az anyagcsere-maradványok és a mérgező anyagok, például a nehézfémek (kadmium, cink és nikkel) megtalálhatók a vakuolákban. Ezenkívül más anyagokat is tárolnak, például pigmenteket (például antocianinokat) a szirmok epidermális sejtjeiben, mérgező anyagokat növényevők ellen, gyantákat, alkaloidokat, például ópiumot stb. A gyümölcsök és zöldségek ízének nagy része a vakuolákban tárolt anyagok eredménye.
degradációs központok
litikus vakuolák találhatók a vegetatív szövetekben, ezért vegetatív vakuoláknak is nevezik őket. Tartalmaznak enzimeket, például proteázokat és nukleázokat, valamint számos fehérjét, amelyek részt vesznek a kórokozók elleni védekezésben. A vakuolumembránba helyezett protonpumpák protonokat juttatnak a vakuolába, és megsavanyítják a belső tartalmat. Az alacsony pH és a litikus enzimek lehetővé teszik a lebomlási folyamatokat. A vakuolok hasonló szerepet játszanak az állati sejtek lizoszómáiban. Továbbá, a lizoszómákhoz hasonlóan, a vakuolok is részt vesznek az autofágiában. A vakuoláris feldolgozó enzimek olyan fehérjék, amelyek a vakuolákban is megtalálhatók. Átalakítják a vakuolába inaktív molekulákként érkező molekuláris prekurzorokat aktív molekulákká.
apoptózis
a Vakuolok autolízis néven ismert mechanizmuson keresztül vesznek részt a növényi sejt apoptózisában. Ezenkívül a túlérzékeny sejthalál néven ismert sejthalál a növényi sejtekben akkor fordul elő, amikor a vakuolamembrán megszakad.
Egyéb
különböző növényi szövetekben speciális vakuolok vannak. Például a mag belső tegumentumaiban a vakuolok felhalmozódnak a flavonoidokban az ultraibolya fény elleni védelem érdekében. A flavonoidokat az endoplazmatikus retikulum membránok citoszolos felületén szintetizálják, majd a vakuolák belsejébe transzlokálják a végső kémiai feldolgozás céljából. A vacuole membránban vannak transzporterek, amelyek elvégzik ezt a transzlokációt.
néhány növényfaj, mint a brassicas, vegetatív szöveteiben vacuolák vannak a növényevők visszaszorítására. Ezek a vakuolok fehérjéket, például myrosinázokat tárolnak. Miután a növényevő aktivitás felszabadult, ezek az enzimek lebontják a levelek molekuláris vegyületeit, amelyek mérgezővé válnak az állat számára. A myrosint tároló sejteket myrosine sejteknek nevezik, és a levelek érrendszeri kötegei közelében találhatók.
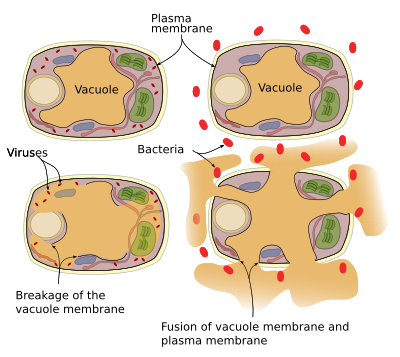
a növényeknek nincs immunrendszerük, így minden sejtnek megvan a saját védelmi rendszere. A védelmi fehérjék és enzimek a vakuolákban találhatók. Két védelmi mechanizmus létezik, amelyeket a vakuolák képesek végrehajtani (3.ábra): a vakuol membrán összeomlik és a membrán vakuol fúziója a plazmamembránnal. A vírusfertőzések vakuol membrántöréshez vezetnek, és enzimeket szabadítanak fel a citoszolba, ahol megtámadhatják a vírusokat. A vakuolamembrán és a plazmamembrán fúziója felszabadítja a vakuolamembrán enzimeket az extracelluláris térbe, ahol megölhetik a kórokozókat, mint a baktériumokat.
hólyagos kereskedelem
a Vakuolok a hólyagos forgalom részét képezik. Valójában a hólyagos kereskedelem végtermékének tekinthetők, mivel kialakulásuk és fenntartásuk a bejövő vezikulumoktól függ. Molekulák, amelyeket tárolni vagy lebontani fognak, beleértve a hidrolitikus enzimeket, valamint az összes membránmolekulát a vakuolákra célozzák vezikulákon keresztül. A molekulák különböző vezikuláris utakat követhetnek a vakuolák eléréséhez:
endoplazmatikus retikulum> Golgi készülék> Vacuole; Golgi készülék> pre-vacuoláris rekesz > vacuole. Ez az alapértelmezett út a hidrolitikus enzimek vacuolák felé történő szállításához. A pre-vacuoláris rekeszek hasonlóak az állati sejtek multivesikuláris testeihez/ késői endoszómáihoz. Kíváncsi, hogy a hidrolitikus enzimeket a Golgi-készülékben nem 6-foszfát-mannóz részek választják ki, hanem az aminosavláncukban található aminosavak szekvenciájával. Vannak specifikus aminosavszekvenciák, amelyek a fehérjéket a hidrolitikus vakuolákhoz irányítják, más szekvenciák pedig specifikusak arra, hogy más fehérjéket a tároló vakuolák felé mozgatjanak. A vakuolákra célzott összes fehérjének van egy jelszekvenciája, és ezeket a receptoroknak specifikusan fel kell ismerniük.
endoplazmatikus retikulum> vakuol. A molekulák közvetlenül az endoplazmatikus retikulumból érkezhetnek a vakuolákba. Ez az út kiemelkedő a magokban, mint tárolási út. Azonban más növényi sejtekben, mint a levelekben, ez az út nagyon ritka lehet. Az endoplazmatikus retikulumból a vacuolákba utazó vezikulák függetlenek a COP-II bevonatoktól, amelyek a Golgi-készülékre célzott vezikulákhoz szükségesek. Az endoplazmatikus retikulum-vakuol útvonalban néha vannak köztes rekeszek, de átmeneti membránhoz kötött organellák, ahol a molekulák rövid időn belül megmaradnak, mielőtt megérkeznek a vakuolába. Ez a vezikuláris út származhat autofágiás sejtkomponensekből.
plazmamembrán > vakuol. Az endocitikus vezikulák közvetlenül vakuolákkal olvadnak össze, amelyek úgy működnek, mint a korai endoszómák.
Bibliográfia
Marty F. 1999. Növényi vakuolok. Növényi sejt 11: 587-600.
Pereira C., Pereira S, Pissarra J. 2014. Fehérjéket szállítunk a növényi vacuolába — egy frissítés. Nemzetközi molekuláris tudományos folyóirat 15: 7611-762.
Simada T, Takagi J, Ichino T, Sirakava M, Hara-Nisimura I. 2018. Növényi vakuola. Éves áttekintés a növénybiológiában. https://doi.org/10.1146/annurev-arplant-042817-040508.
Taiz L. 1992. A növény vakuolája. Kísérleti biológiai folyóirat 172: 113-122.
Zhang C, Hicks G R, Raikhel NV. 2014. Növényi vacuole morfológia és vacuolaris kereskedelem. Határok a növénytudományokban 5: 476. ![]()