1. Ominaisuudet
2. Funktiot
– Turgor
– Varastointi
– Lyytinen
– Apoptoosi
– Muut
3. Vesikulaarikauppa
Vakuolit ovat kalvoon sitoutuneita organelleja, joita esiintyy kasvisoluissa ja sienissä, myös hiivoissa. Ne ovat kasvisolujen toiminnan kannalta kriittisiä organelleja.
ominaisuudet
Vakuolit ovat yleensä suuria osastoja, joita täysikasvuisissa soluissa voi olla jopa 90% solun kokonaistilavuudesta (Kuvat 1 ja 2). Ne ovat kasvisolujen suurin osasto. Nimi vacuole on johdettu latinan sanasta” vacuuus”, joka tarkoittaa tyhjää. Tämä oli selvästi väärinkäsitys, koska vakuolit eivät ole tyhjiä, vaan täynnä enemmän tai vähemmän väkevää vesiliuosta. Vakuolin kalvo tunnetaan tonoplastina, ja se on olennainen osa tämän organellen toimintaa. Kasveissa on useita erilaisia vakuoleja niiden suorittaman roolin mukaan. Kasvisolu voi sisältää erityyppisiä vakuoleja, ja vakuoli voi muuttaa entsyymivalikoimaansa ja sitten muuttaa toimintaansa.
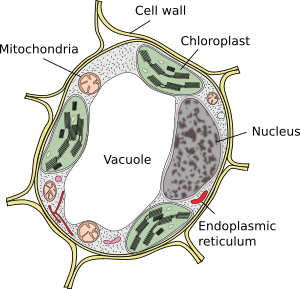
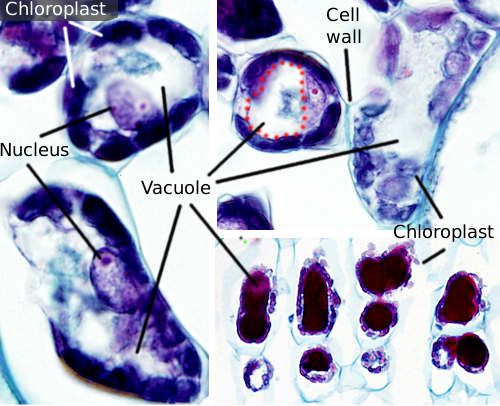
Vakuolit ovat yleensä pyöristettyjä, mutta lopulliseen muotoon vaikuttaa solujen morfologia. Yksi suuri vakuoli havaitaan usein kypsissä kasvisoluissa. Vakuolin kalvo kuitenkin taittuu joskus syvälle ja vuolaasti ja muodostaa pieniä lokeroita, jotka näyttävät monilta pieniltä vakuoleilta valomikroskopiassa havaittaessa, mutta ne ovat todellisuudessa vain yksi vakuoli, koska kalvo on jatkuva.
uusia vakuoleja muodostuu Golgin laitteesta vapautuneiden vesikkelien fuusioituessa. Aluksi ne muodostavat uuden osaston, joka tunnetaan nimellä pro-vacuole. Meristemaattisessa solussa voi olla satoja pro-vakuoleja. Sitten solujen erilaistumisen aikana pro-vacuolit sulautuvat keskenään pieniksi vakuoleiksi, ja fuusioprosessi jatkuu, kunnes muodostuu suuri keskusvakuoli. Endoplasmainen verkkokalvo saattaa osallistua myös vakuolien muodostumiseen ja kasvuun joissakin kasvisoluissa, lähinnä siemenissä. Kun suuri vakuoli on läsnä, Golgin laitteen ja plasmakalvon vesikkelit säätelevät kokoa lisäämällä ja poistamalla kalvoa.
useimpien kasvisolujen tärkein vakuoli on suuri osasto, joka on täytetty happamalla liuoksella, joka sisältää suoloja (natrium, kalium), aineenvaihduntatuotteita (hiilihydraatit, orgaaniset hapot) ja joitakin pigmenttejä. Osa näistä molekyyleistä pääsee vakuoliin sytosolista pitoisuusgradienttia vastaan. Normaali pH vakuolin sisällä vaihtelee välillä 5 ja 5,5, vaikka se voi olla sitruunahedelmässä noin 2 tai joillakin Levillä jopa 0,6.
funktio
Vakuolit ovat välttämättömiä kasvisolujen fysiologialle ja homeostaasille, ja ne suorittavat erilaisia tehtäviä solutyypin mukaan.
Turgor
Cell turgor on hydrostaattisen paineen taso kasvisolun soluseinää vastaan. Tätä painetta valvovat vakuolit, jotka saavat sisäänsä erilaisia aineita, kuten ioneja, tuottaakseen muuttuvia sisäisiä osmoottisia ympäristöjä verrattuna sytosolin ympäristöihin. Vakuolikalvon molemmin puolin oleva erilainen osmolaarisuus saa veden kulkemaan kalvon poikki joko sisäänpäin tai ulospäin. Tyhjiön osmolaarisuuteen osallistuvat aineet voivat ylittää vakuolikalvon ionipumppujen välittämän ATP-riippuvaisen kuljetuksen avulla. H (+)-ATPaasi ja H (+)-pyrofosfataasi kykenevät muodostamaan protonigradientteja vakuolikalvon molempien sivujen välille, ja näitä gradientteja käytetään muiden molekyylien kuljettamiseen. Kyky varastoida vettä vakuolin sisään on välttämätöntä kasvien solujen kasvulle mitoosin jälkeen. Kasvisolut voivat kasvattaa kokoaan 10-20-kertaiseksi, mikä on erittäin hyödyllistä ruumiskasvin kasvulle ja kasvien elinten muodon muokkaamiselle. Hydrostaattisen paineen välittämä kasvu säästää energiaa, koska on halvempaa lisätä veden määrää kuin syntetisoida uusia molekyylejä (eläinsolujen kasvu perustuu molekyylisynteesiin). Kasvisolujen on turvallisempaa kerryttää vettä vakuoliin, koska näin sytosolumolekyylit eivät laimene, mikä vaarantaisi solujen selviytymisen.
varastointi
Vakuolit ovat viimeinen asema joillekin vesicular – liikenneväylille. Joissakin soluissa ne ovat hiilihydraattien ja proteiinien säilytyslokero. Tämä tapahtuu selvästi siemenissä, joissa vakuoleihin kertyy itämisen aikana tarvittavia proteiineja. Varastointi vakuolit muuttuvat lytic vakuolit aikana solujen erilaistuminen. Toisin kuin eläimillä, kasveilla ei ole erittymisjärjestelmää, eivätkä ne voi liikkua myrkyllisten aineiden välttämiseksi. Kasveissa mahdollisesti vaarallisia aineita varastoidaan vakuoleihin. Näin vakuoleista löytyy aineenvaihdunnallisia jäämiä ja myrkyllisiä aineita, kuten raskasmetalleja (kadmiumia, sinkkiä ja nikkeliä). Lisäksi ne varastoivat myös muita aineita, kuten pigmenttejä (esimerkiksi antosyaaneja) terälehtien epidermaalisiin soluihin, myrkyllisiä aineita kasvinsyöjiä vastaan, hartseja, alkaloideja kuten oopiumia jne. Suurin osa hedelmien ja vihannesten mausta johtuu vakuoleihin varastoiduista aineista.
Hajoamiskeskuksia
lyyttisiä vakuoleja löytyy vegetatiivisista kudoksista, joten niitä kutsutaan myös vegetatiivisiksi vakuoleiksi. Ne sisältävät entsyymejä, kuten proteaaseja ja nukleaaseja, sekä useita proteiineja, jotka osallistuvat puolustukseen taudinaiheuttajia vastaan. Vakuolikalvoon työnnetyt protonipumput syöttävät vakuoliin protoneja ja happamoittavat sisätilojen sisältöä. Alhainen pH ja lyyttiset entsyymit mahdollistavat hajoamisprosessit. Vakuoleilla on samanlainen rooli kuin eläinsolujen lysosomeilla. Lisäksi tyhjiöt osallistuvat lysosomien tavoin autofagiaan. Vacuolar processing entsyymit ovat proteiineja, joita esiintyy myös vakuoleissa. Ne muuttavat vakuoliin inaktiivisina molekyyleinä saapuvat molekyylien esiasteet aktiivisiksi molekyyleiksi.
apoptoosi
Vakuolit osallistuvat kasvisolujen apoptoosiin autolyysiksi kutsutun mekanismin kautta. Lisäksi kasvin soluissa tapahtuu yliherkäksi solukuolemaksi kutsuttu solukuolema, kun vakuolikalvo rikkoutuu.
muut
eri kasvikudoksissa on erikoistuneita vakuoleja. Esimerkiksi siemenen sisäisiin tegumentteihin vakuolit keräävät flavonoideja suojaamaan ultraviolettivalolta. Flavonoideja syntetisoidaan solulimakalvoston solupinnalla ja siirretään sitten vakuolien sisäpuolelle lopullista kemiallista käsittelyä varten. Vakuolikalvossa on kuljettajia, jotka suorittavat tämän translokaation.
joillakin kasvilajeilla, kuten brassicoilla, on kasvullisissa kudoksissaan vakuoleja kasvinsyöjien kumoamiseksi. Nämä vakuolit varastoivat proteiineja, kuten myrosinaaseja. Kun kasvinsyöjäaktiivisuus vapauttaa nämä entsyymit, ne hajottavat lehtien molekyyliyhdisteitä, joista tulee myrkyllisiä eläimelle. Myrosiinia varastoivia soluja kutsutaan myrosiinisoluiksi ja niitä löytyy läheltä lehtien verisuonikimppuja.
kasveilta puuttuu immuunijärjestelmä, joten jokaisella solulla on oma puolustusjärjestelmänsä. Puolustusproteiineja ja entsyymejä löytyy vakuoleista. Vakuoleilla voidaan suorittaa kaksi puolustusmekanismia (kuva 3): vakuolikalvo romahtaa ja kalvon vakuoli fuusioituu plasmakalvoon. Virusinfektiot johtavat vakuolikalvon rikkoutumiseen ja vapauttavat entsyymejä sytosoliin, jossa ne voivat hyökätä viruksia vastaan. Vakuolikalvon ja plasmakalvon fuusio vapauttaa vakuolientsyymejä solunulkoiseen tilaan, jossa ne voivat tappaa taudinaiheuttajia, kuten bakteereja.
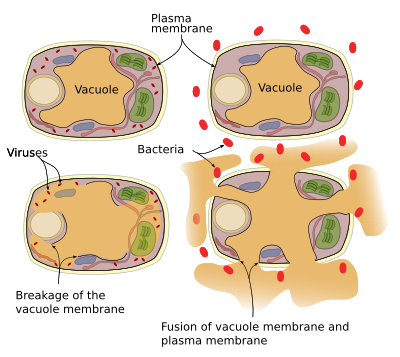
Vesicular traffic
Vacuolit ovat osa vesicular-liikennettä. Itse asiassa niitä voidaan pitää vesikulaarikaupan lopputuotteena, koska niiden muodostuminen ja säilyminen riippuu saapuvista vesikkeleistä. Varastoitavat tai hajoavat molekyylit, mukaan lukien hydrolyyttiset entsyymit, sekä kaikki kalvomolekyylit ohjataan vesikkelien kautta vakuoleihin. Molekyylit voivat seurata eri vesikulaarireittejä päästäkseen vakuoleihin:
Endoplasminen retikulaatti> Golgin laite> vakuoli; Golgin laite> Pre-vakuoliosasto > vakuoli. Tämä on oletusreitti hydrolyyttisten entsyymien kuljettamiseksi kohti vakuoleja. Pre-vacuolar lokerot ovat samanlaisia kuin monivivahteiset kappaleet / eläinsolujen myöhäiset endosomit. Kummallista kyllä, hydrolyyttisiä entsyymejä ei valita Golgin laitteessa 6-fosfaatti-mannoosiosuuksilla, vaan niiden aminohappoketjussa sijaitsevien aminohappojen sekvenssillä. On olemassa erityisiä sekvenssejä aminohappoja kohdentamiseksi proteiineja hydrolyyttisiä vakuoleja ja muut sekvenssit ovat erityisiä siirtää muita proteiineja kohti varastoiva vakuoleja. Kaikilla vakuoleihin kohdistetuilla proteiineilla on signaalijakso, ja ne täytyy tunnistaa nimenomaan reseptoreista.
endoplasmainen retikulaatti> vakuoli. Molekyylit voivat päätyä vakuoleihin suoraan endoplasmaisesta retikulaatista. Tämä reitti on merkittävä siemenissä varastointireittinä. Muissa kasvisoluissa, kuten lehdissä, tämä reitti saattaa kuitenkin olla hyvin harvinainen. Endoplasmaisesta retikulumista vakuoleihin kulkevat vesikkelit ovat riippumattomia COP-II-päällysteistä, joita tarvitaan Golgin laitteeseen kohdistettuihin vesikkeleihin. Endoplasmaisessa retikulum-vakuolireitissä on joskus väliosia, mutta ne ovat ohimeneviä kalvoon sitoutuneita organelleja, joissa molekyylit säilyvät vähän ennen kuin ne saapuvat vakuoliin. Tämä vesikulaarireitti voi olla peräisin autofagisista solukomponenteista.
Plasmakalvo > vakuoli. Endosyyttiset vesikkelit sulautuvat suoraan vakuoleihin, jotka toimivat kuten varhaiset endosomit.
Bibliografia
Marty F. 1999. Istuta vakuoleja. Kasvisolu 11: 587-600.
Pereira C., Pereira S, Pissarra J. 2014. Proteiinien toimittaminen vakuoliin. päivitys. International journal of molecular sciences 15: 7611-762.
Shimada T, Takagi J, Ichino T, Shirakawa M, Hara-Nishimura I. 2018. Istuta vakuoli. Vuosikatsaus kasvibiologiassa. https://doi.org/10.1146/annurev-arplant-042817-040508.
Taiz L. 1992. Kasvin vakuoli. Journal of experimental biology 172: 113-122.
Zhang C, Hicks G R, Raikhel NV. 2014. Plant vacuole morphology ja vacuolar traffic. Frontiers in plant sciences 5: 476. ![]()