1. Kenmerken
2. Functies
– Turgor
– Opslag
– Lytische
– Apoptose
– Overige
3. Vesiculaire handel
vacuolen zijn membraangebonden organellen die in plantencellen en schimmels, met inbegrip van gisten, worden aangetroffen. Zij zijn kritieke organellen voor de functie van de plantaardige cel.
features
vacuolen zijn meestal grote compartimenten die in volwassen cellen tot 90% van het totale celvolume kunnen bedragen (figuren 1 en 2). Ze zijn het grootste compartiment van plantencellen. De naam vacuole is afgeleid van het Latijnse woord “vacuus”, wat leeg betekent. Dit was duidelijk een misverstand omdat vacuolen niet leeg zijn, maar gevuld met een min of meer geconcentreerde waterige oplossing. Het membraan van de vacuole staat bekend als tonoplast, en het is een essentieel onderdeel voor de functie van deze organelle. In planten zijn er verschillende soorten vacuolen volgens de rol die ze uitvoeren. Een installatiecel kan verschillende soorten vacuolen bevatten, en een vacuole kan zijn enzymrepertoire wijzigen en dan zijn functie veranderen.
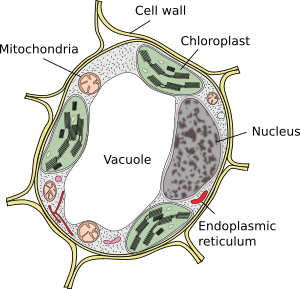
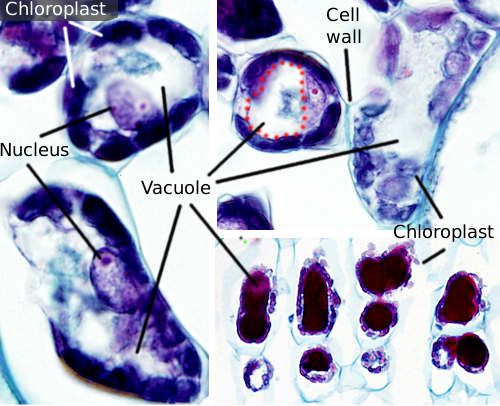
vacuolen zijn meestal afgerond, maar de uiteindelijke vorm wordt beïnvloed door celmorfologie. Een grote vacuole wordt vaak waargenomen in volwassen plantencellen. Echter, het membraan van de vacuole wordt soms diep en overvloedig gevouwen en vormt kleine compartimenten die eruit zien als vele kleine vacuole wanneer waargenomen bij lichtmicroscopie, maar ze zijn eigenlijk slechts één vacuole omdat membraan continu is.
nieuwe vacuolen worden gevormd door fusie van blaasjes die vrijkomen uit het Golgi-apparaat. Aanvankelijk vormen ze een nieuw compartiment dat bekend staat als pro-vacuole. Een meristematische cel kan honderden Pro-vacuolen hebben. Dan, tijdens celdifferentiatie, fuseren pro-vacuolen tussen elkaar in kleine vacuolen, en het fusieproces gaat door totdat een grote centrale vacuole wordt gevormd. Het endoplasmatisch reticulum kan ook betrokken zijn bij de vorming en groei van vacuolen in sommige plantencellen, meestal in zaden. Zodra een grote vacuole aanwezig is, regelen blaasjes van het Golgi-apparaat en plasmamembraan de grootte door membraan toe te voegen en te verwijderen.
de belangrijkste vacuole van de meeste plantencellen is een groot compartiment gevuld met een zure oplossing die zouten (natrium, kalium), metabolieten (koolhydraten, organische zuren) en enkele pigmenten bevat. Sommige van deze molecules gaan vacuole van cytosol tegen concentratiegradiënt in. De normale pH in de vacuole varieert tussen 5 en 5,5, hoewel het ongeveer 2 kan zijn in de citroenfruit, of zelfs 0,6 in sommige algen.
functie
vacuolen zijn essentieel voor de fysiologie en homeostase van plantencellen en vervullen verschillende functies naargelang het celtype. Hieronder volgen enkele voorbeelden:
Turgor
cel turgor is het niveau van hydrostatische druk tegen de celwand van de plantcel. Deze druk is onder de controle van vacuolen, die verschillende substanties binnen, met inbegrip van ionen krijgen, om veranderlijke binnen osmotic milieu ‘ s te veroorzaken wanneer vergeleken met die van cytosol. De verschillende osmolariteit aan beide zijden van het vacuole membraan zorgt ervoor dat het water het membraan kruist, naar binnen of naar buiten. De stoffen die bijdragen aan de vacuole osmolariteit kunnen het vacuole membraan passeren door ATP afhankelijk transport gemedieerd door ionische pompen. H(+)-ATPase en H ( + ) – pyrofosfatase kunnen protongradiënten tussen beide kanten van het vacuole membraan vormen, en deze gradiënten worden gebruikt om andere molecules te vervoeren. Het vermogen om water in de vacuole op te slaan is essentieel voor de groei van plantencellen na mitose. Plantencellen kunnen hun grootte 10 tot 20 keer vergroten, wat erg handig is voor de lichaamsplant om te groeien en voor het wijzigen van de vorm van plantenorganen. De groei gemedieerd door hydrostatische druk bespaart energie omdat het goedkoper is om de hoeveelheid water te verhogen dan het synthetiseren van nieuwe moleculen (dierlijke celgroei is gebaseerd op moleculaire synthese). Het is veiliger voor plantencellen om water te accumuleren in de vacuole omdat op deze manier de cytosolic moleculen niet verdund worden, wat de celoverleving in gevaar zou brengen.
opslag
vacuolen zijn het laatste station voor sommige vesiculaire verkeerswegen. In sommige cellen zijn ze het compartiment om koolhydraten en eiwitten op te slaan. Dit gebeurt duidelijk in zaden, waar vacuolen eiwitten accumuleren die nodig zijn tijdens de ontkieming. Opslag vacuolen worden lytische vacuolen tijdens celdifferentiatie. In tegenstelling tot dieren hebben planten geen uitscheidingssysteem, noch kunnen ze zich verplaatsen om giftige stoffen te vermijden. In installaties worden potentieel gevaarlijke stoffen opgeslagen in vacuolen. Op deze manier worden stofwisselingsresiduen en giftige stoffen zoals zware metalen (cadmium, zink en nikkel) in vacuolen aangetroffen. Daarnaast slaan ze ook andere stoffen op zoals pigmenten (bijvoorbeeld anthocyanen) in de epidermale cellen van bloemblaadjes, giftige stoffen tegen herbivoren, harsen, alkaloïden zoals opium, enzovoort. Het grootste deel van de smaak van fruit en groenten is het resultaat van stoffen opgeslagen in vacuolen.
Afbraakcentra
lytische vacuolen kunnen worden gevonden in vegetatieve weefsels, zodat ze ook bekend staan als vegetatieve vacuolen. Zij bevatten enzymen zoals proteasen en nucleases, evenals een aantal proteã nen betrokken bij de verdediging tegen ziekteverwekkers. Protonpompen die in het vacuole membraan zijn ingebracht, voeren protonen in de vacuole in en verzuren de inwendige inhoud. De lage pH en de lytische enzymen staan afbraakprocessen toe. Vacuolen hebben een gelijkaardige rol aan lysosomen van dierlijke cellen. Verder, zoals lysosomes, nemen vacuoles in autophagy deel. Vacuolaire verwerkingsenzymen zijn proteã nen ook gevonden in vacuolen. Ze transformeren moleculaire precursoren die in de vacuole aankomen als inactieve moleculen in actieve moleculen.
apoptose
vacuolen zijn betrokken bij plantcelapoptose via een mechanisme dat autolyse wordt genoemd. Bovendien, komt een type van celdood bekend als overgevoelige celdood voor in installatiecellen wanneer het vacuolemembraan wordt gebroken.
andere
er zijn gespecialiseerde vacuolen in verschillende plantenweefsels. Bijvoorbeeld, in het zaad interne tegumenten, vacuolen accumuleren flavonoïden voor bescherming tegen ultraviolet licht. Flavonoïden worden gesynthetiseerd in het cytosolisch oppervlak van de endoplasmatische reticulummembranen en vervolgens getransloceerd naar het interieur van vacuolen voor een definitieve chemische verwerking. In het vacuole membraan zijn er transporters om deze translocatie uit te voeren.
sommige plantensoorten, zoals Brassica ‘ s, hebben vacuolen in hun vegetatieve weefsels voor het verwijderen van herbivoren. Deze vacuolen slaan eiwitten op, zoals myrosinases. Eenmaal vrijgegeven door de herbivore activiteit, deze enzymen degraderen moleculaire verbindingen van de bladeren die giftig voor het dier. De cellen die myrosine opslaan staan bekend als myrosinecellen en kunnen dichtbij de vasculaire bundels van bladeren worden gevonden.
planten hebben geen afweersysteem, zodat elke cel zijn eigen afweersysteem heeft. De proteïnen en enzymen van de verdediging kunnen in vacuolen worden gevonden. Er zijn twee afweermechanismen die vacuolen kunnen uitvoeren (Figuur 3): vacuole membraan instort en fusie van membraan vacuole met plasma membraan. De virale besmettingen leiden tot vacuole membraanbreuk en geven enzymen in cytosol vrij, waar zij virussen kunnen aanvallen. De fusie van vacuole membraan en plasmamembraan geeft vacuole enzymen aan de extracellulaire ruimte vrij waar zij ziekteverwekkers zoals bacteriën kunnen doden.
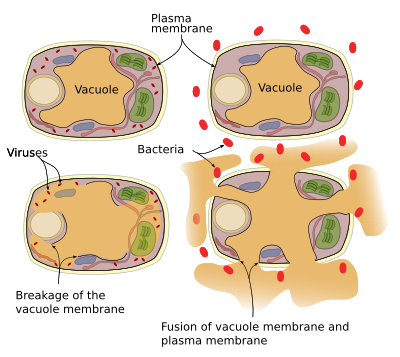
vesiculaire handel
vacuolen maken deel uit van het vesiculaire verkeer. Eigenlijk kunnen ze worden beschouwd als een eindproduct van de vesiculaire handel omdat hun vorming en onderhoud afhankelijk is van de inkomende blaasjes. De Molecules die zullen worden opgeslagen of gedegradeerd, inbegrepen hydrolytische enzymen, evenals alle membraanmolecules worden gericht aan vacuolen via blaasjes. Moleculen kunnen verschillende vesiculaire routes volgen om vacuolen te bereiken:
endoplasmatisch reticulum> Golgi-apparaat> Vacuol; Golgi-apparaat> pre-vacuolair compartiment > vacuol. Dit is de standaardweg om hydrolytische enzymen naar vacuolen te transporteren. De pre-vacuolaire compartimenten zijn gelijkaardig aan multivesicular lichamen / late endosomes van dierlijke cellen. Vreemd genoeg worden hydrolytische enzymen in het Golgi-apparaat niet geselecteerd door 6-fosfaat-mannose-delen, maar door een opeenvolging van aminozuren in hun aminozuurketen. Er zijn specifieke opeenvolgingen van aminozuren voor het richten van proteã nen aan hydrolytic vacuoles en andere opeenvolgingen zijn specifiek voor het bewegen van andere proteã nen naar het opslaan vacuoles. Alle proteã nen die aan vacuoles worden gericht hebben een signaalopeenvolging, en zij moeten specifiek door receptoren worden erkend.
endoplasmatisch reticulum> vacuole. De Molecules kunnen aan vacuolen direct van het endoplasmic reticulum aankomen. Deze weg is prominent in zaden als weg voor opslag. Echter, in andere plantencellen, zoals in bladeren, kan dit pad zeer zeldzaam zijn. Blaasjes die van het endoplasmatisch reticulum naar vacuolen reizen, zijn onafhankelijk van Cop-II vachten, die nodig zijn voor blaasjes gericht op het Golgi-apparaat. In de endoplasmic reticulum-vacuole weg, zijn er soms tussencompartimenten, maar zij zijn voorbijgaande membraan-gebonden organellen waar de molecules kort worden behouden alvorens zij aan de vacuole aankomen. Deze vesiculaire weg kan uit autophagy cellulaire componenten worden afgeleid.
plasmamembraan > vacuole. Endocytische blaasjes smelten direct samen met vacuolen, die als vroege endosomen werken.
Bibliografie
Marty F. 1999. Plaats vacuolen. Plant cell 11: 587-600.
Pereira C., Pereira S, Pissarra J. 2014. Leveren van eiwitten aan de plant vacuole-een update. International journal of molecular sciences 15: 7611-762.
Shimada T, Takagi J, Ichino T, Shirakawa M, Hara-Nishimura I. 2018. Plant vacuole. Jaaroverzicht in plantenbiologie. https://doi.org/10.1146/annurev-arplant-042817-040508.
Taiz L. 1992. De vacuole van de fabriek. Journal of experimental biology 172: 113-122.
Zhang C, Hicks G R, Raikhel NV. 2014. Plant vacuole morfologie en vacuolaire handel. Grenzen in plantenwetenschappen 5: 476. ![]()