1. Características
2. Funções
– Turgor
– Armazenamento
– Letic
– Apoptose
– Outros
3. Tráfico Vesicular
vacúolos são organelos ligados à membrana encontrados em células vegetais e fungos, incluindo leveduras. São organelas críticas para a função celular das plantas.
características
vacúolos são geralmente grandes compartimentos que em células maduras podem ser até 90% do volume total de células (Figuras 1 e 2). Eles são o maior compartimento de células vegetais. O nome vacuole é derivado da palavra latina vacuus, que significa vazio. Isto foi claramente um mal-entendido porque os vacúolos não estão vazios, mas preenchidos com uma solução aquosa mais ou menos concentrada. A membrana do vacúolo é conhecida como tonoplasto, e é uma parte essencial para a função desta organela. Nas plantas, existem vários tipos de vacúolos de acordo com o papel que desempenham. Uma célula vegetal pode conter diferentes tipos de vacúolos, e um vacúolo pode modificar seu repertório enzimático e, em seguida, mudar sua função.
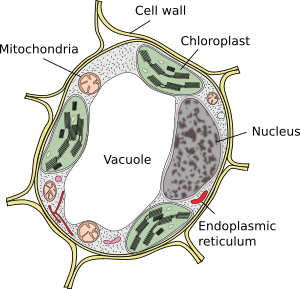
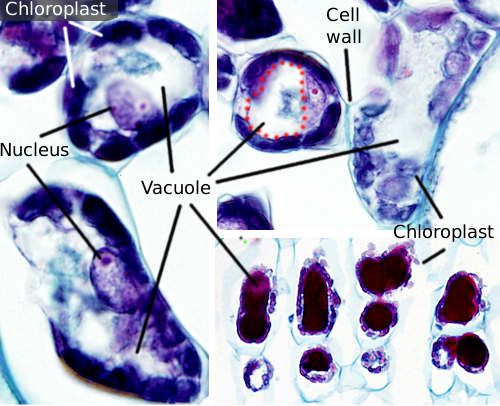
novos vacúolos são formados pela fusão das vesículas libertadas do aparelho de Golgi. Inicialmente formam um novo compartimento conhecido como pro-vacuole. Uma célula meristemática pode ter centenas de pró-vacúolos. Então, durante a diferenciação celular, pro-vacúolos se fundem entre si em pequenos vacúolos, e o processo de fusão continua até que um grande vacúolo central é formado. O retículo endoplásmico também pode estar envolvido na formação e crescimento de vacúolos em algumas células vegetais, principalmente em sementes. Uma vez que um grande vacúolo está presente, vesículas do aparelho de Golgi e membrana plasmática regulam o tamanho, adicionando e removendo membrana.
o principal vacuole da maioria das células vegetais é um grande compartimento cheio de uma solução ácida contendo sais (sódio, potássio), metabolitos (hidratos de carbono, ácidos orgânicos) e alguns pigmentos. Algumas destas moléculas entram no vácuo do citosol contra o gradiente de concentração. O pH normal dentro do vacúolo varia entre 5 e 5,5, embora possa ser cerca de 2 no fruto de limão, ou mesmo 0,6 em algumas algas.
função
vacúolos são essenciais para a fisiologia e homeostase das células vegetais, e desempenham funções diferentes de acordo com o tipo celular. Alguns deles são:
Turgor
turgor celular é o nível de pressão hidrostática contra a parede celular da célula vegetal. Esta pressão está sob o controle de vacúolos, que colocam diferentes substâncias no interior, incluindo íons, para produzir ambientes osmóticos internos variáveis quando comparados com os do citosol. A osmolaridade diferente em ambos os lados da membrana vacúola faz com que a água atravesse a membrana, tanto para dentro como para fora. As substâncias que contribuem para a osmolaridade do vacúolo podem atravessar a membrana do vacúolo por transporte dependente de ATP mediado por bombas iónicas. H (+)-ATPase e H (+)-pirofosfatase são capazes de formar gradientes de prótons entre ambos os lados da membrana vacuole, e estes gradientes são usados para transportar outras moléculas. A capacidade de armazenar água dentro do vacuole é essencial para o crescimento de células vegetais após a mitose. As células da planta podem aumentar seu tamanho 10 a 20 vezes, o que é muito útil para a planta do corpo crescer e para modificar a forma dos órgãos da planta. O crescimento mediado pela pressão hidrostática economiza energia porque é mais barato aumentar a quantidade de água do que sintetizar novas moléculas (o crescimento de células animais é baseado na síntese molecular). É mais seguro para as células das plantas acumularem água no vacúolo porque desta forma as moléculas citosólicas não se diluem, o que comprometeria a sobrevivência celular.
Armazenamento
vacúolos são a última estação para algumas vias de tráfego vesicular. Em algumas células, elas são o compartimento para armazenar carboidratos e proteínas. Isto acontece claramente em sementes, onde vacúolos acumulam proteínas necessárias durante a germinação. Vacúolos de armazenamento tornam-se vacúolos líticos durante a diferenciação celular. Ao contrário dos animais, as plantas não têm um sistema de excreção, nem podem mover-se para evitar substâncias tóxicas. Nas plantas, as substâncias potencialmente perigosas são armazenadas em vacúolos. Desta forma, resíduos do metabolismo e substâncias tóxicas como metais pesados (cádmio, zinco e níquel) são encontrados em vacúolos. Além disso, também armazenam outras substâncias como pigmentos (por exemplo, antocianinas) nas células epidérmicas de pétalas, substâncias tóxicas contra herbívoros, resinas, alcalóides como o ópio, etc. A maior parte do sabor das frutas e legumes é o resultado de substâncias armazenadas em vacúolos.
Centros de degradação
vacúolos líticos podem ser encontrados em tecidos vegetativos, por isso também são conhecidos como vacúolos vegetativos. Eles contêm enzimas como proteases e nucleases, bem como uma série de proteínas envolvidas na defesa contra patógenos. Bombas de protões inseridas na membrana vacuole entram protões no vacuole e acidificam o conteúdo interior. O baixo pH e as enzimas líticas permitem processos de degradação. Vacúolos têm um papel semelhante aos lisossomas das células animais. Além disso, como lisossomas, vacúolos participam na autofagia. Enzimas de processamento vacuolares são proteínas também encontradas em vacúolos. Transformam precursores moleculares que chegam ao vácuo como moléculas inactivas em moléculas activas.
apoptose
vacúolos estão envolvidos na apoptose das células vegetais através de um mecanismo conhecido como autólise. Além disso, um tipo de morte celular conhecida como morte celular hipersensível ocorre nas células vegetais quando a membrana do vacúolo é quebrada.
outros
existem vacúolos especializados em diferentes tecidos vegetais. Por exemplo, nos tegumentos internos das sementes, os vacúolos acumulam flavonóides para proteção contra a luz ultravioleta. Os flavonóides são sintetizados na superfície citosólica das membranas reticulum endoplasmáticas e, em seguida, translocados para o interior dos vacúolos para um processamento químico final. Na membrana vacúola há transportadores para realizar esta translocação. Algumas espécies de plantas, como brassicas, têm vacúolos em seus tecidos vegetativos para revogar herbívoros. Estes vacúolos armazenam proteínas, como as mirosinases. Uma vez libertadas pela actividade herbívora, estas enzimas degradam compostos moleculares das folhas que se tornam tóxicas para o animal. As células que armazenam mirosina são conhecidas como células mirosina e podem ser encontradas perto dos feixes vasculares das folhas.
as plantas não têm sistema imunitário, de modo que cada célula tem o seu próprio sistema de defesa. Proteínas de defesa e enzimas podem ser encontradas em vacúolos. Existem dois mecanismos de defesa que vacúolos podem realizar (Figura 3): a membrana vacúola colapsa e fusão do vacúolo de membrana com a membrana plasmática. As infecções virais levam à ruptura da membrana vacúola e libertam enzimas no citosol, onde podem atacar vírus. A fusão da membrana vacuole e da membrana plasmática libera enzimas vacuolares para o espaço extracelular onde podem matar patógenos como bactérias.
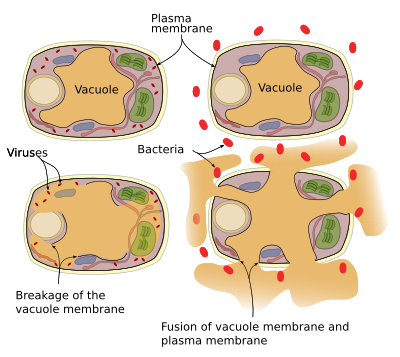
tráfico Vesicular
vacúolos fazem parte do tráfego vesicular. Na verdade, podem ser considerados como um produto final do tráfico vesiculoso, uma vez que a sua formação e manutenção depende das vesículas que chegam. Moléculas que vão ser armazenadas ou degradadas, incluindo enzimas hidrolíticas, bem como todas as moléculas de membrana são direcionadas para vacúolos através de vesículas. As moléculas podem seguir diferentes vesicular caminhos para chegar ao vacúolos:
retículo endoplasmático> o aparelho de Golgi> Vacúolo; o aparelho de Golgi> pré-vacuolar compartimento > vacúolo. Esta é a via padrão para transportar enzimas hidrolíticas para os vacúolos. Os compartimentos pré-vacuolares são semelhantes aos corpos multivesiculares/ endossomas tardios das células animais. Curiosamente, as enzimas hidrolíticas não são selecionadas no aparelho de Golgi por moléculas de 6-fosfato-manose, mas por uma sequência de aminoácidos localizados em sua cadeia de aminoácidos. Existem sequências específicas de aminoácidos para direcionar proteínas para os vacúolos hidrolíticos e outras sequências são específicas para mover outras proteínas para os vacúolos armazenados. Todas as proteínas direccionadas para os vacúolos têm uma sequência de sinal, e precisam de ser reconhecidas especificamente pelos receptores.
retículo endoplasmático > vacuole. Moléculas podem chegar a vacúolos diretamente do retículo endoplasmático. Este caminho é proeminente nas sementes como um caminho para armazenar. No entanto, em outras células vegetais, como nas folhas, esta via pode ser muito rara. Vesículas que viajam do retículo endoplasmático para vacúolos são independentes de COP-II casacos, que são necessários para vesículas direcionadas para o aparelho Golgi. Na Via reticulum-vacuole endoplásmica, existem por vezes compartimentos intermediários, mas são organelas transitórias ligadas à membrana onde as moléculas são retidas pouco antes de chegarem ao vacuolo. Esta via vesicular pode ser derivada de componentes celulares autofágicos.
membrana plasmática > vacuole. As vesículas endocíticas fundem-se directamente com os vacúolos, que funcionam como endossomas iniciais.
Bibliografia
Marty F. 1999. Aspiradores de plantas. Plant cell 11: 587-600.Pereira C., Pereira S, Pissarra J. 2014. Entrega de proteínas ao vacúolo da planta … uma actualização. International journal of molecular sciences 15: 7611-762.
Shimada T, Takagi J, Ichino T, Shirakawa M, Hara-Nishimura I. 2018. Aspirador de planta. Annual review in plant biology. https://doi.org/10.1146/annurev-arplant-042817-040508.
Taiz L. 1992. O buraco de vácuo da planta. Journal of experimental biology 172: 113-122.
Zhang C, Hicks G R, Raikhel NV. 2014. Morfologia de vacuole vegetal e tráfico de vacuolares. Frontiers in plant sciences 5: 476. 