1. Funktioner
2. Funktioner
– Turgor
– Lagring
– Lytisk
– Apoptos
– Andra
3. Vesikulär handel
vakuoler är membranbundna organeller som finns i växtceller och svampar, inklusive jäst. De är kritiska organeller för växtcellsfunktion.
funktioner
vakuoler är vanligtvis stora fack som i mogna celler kan vara upp till 90% av den totala cellvolymen (figurerna 1 och 2). De är det största facket av växtceller. Namnet vacuole härstammar från det latinska ordet ”vacuus”, vilket betyder tomt. Detta var helt klart ett missförstånd eftersom vakuoler inte är tomma utan fyllda med en mer eller mindre koncentrerad vattenlösning. Vakuolmembranet är känt som tonoplast, och det är en väsentlig del för denna organells funktion. I växter finns det flera typer av vakuoler beroende på vilken roll de utför. En växtcell kan innehålla olika typer av vakuoler, och en vakuol kan modifiera sitt enzymrepertoar och sedan ändra dess funktion.
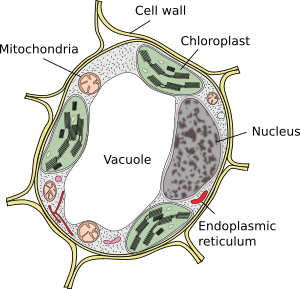
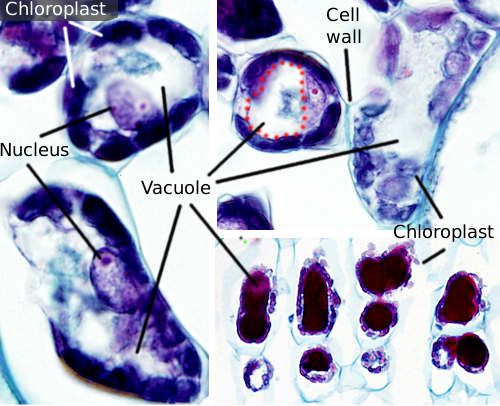
vakuoler är vanligtvis rundade, men den slutliga formen påverkas av cellmorfologi. En stor vakuol observeras ofta i mogna växtceller. Emellertid blir membranet i vakuolen ibland djupt och kraftigt vikta och bildar små fack som ser ut som många små vakuoler när de observeras vid ljusmikroskopi, men de är faktiskt bara en vakuol eftersom membranet är kontinuerligt.
nya vakuoler bildas genom fusion av vesiklar som frigörs från Golgi-apparaten. Ursprungligen bildar de ett nytt fack som kallas pro-vacuole. En meristematisk cell kan ha hundratals Pro-vakuoler. Sedan, under celldifferentiering, smälter Pro-vakuoler mellan varandra i små vakuoler, och fusionsprocessen fortsätter tills en stor central vakuol bildas. Endoplasmatisk retikulum kan också vara involverad i bildandet och tillväxten av vakuoler i vissa växtceller, mestadels i frön. När en stor vakuol är närvarande reglerar vesiklar från Golgi-apparaten och plasmamembranet storleken genom att tillsätta och ta bort membran.
huvudvakuolen hos de flesta växtceller är ett stort fack fyllt med en sur lösning innehållande salter (natrium, kalium), metaboliter (kolhydrater, organiska syror) och vissa pigment. Några av dessa molekyler kommer in i vakuolen från cytosolen mot koncentrationsgradient. Det normala pH-värdet inuti vakuolen varierar mellan 5 och 5,5, även om det kan vara cirka 2 i citronfrukten, eller till och med 0,6 i vissa alger.
funktion
vakuoler är väsentliga för fysiologi och homeostas av växtceller och utför olika funktioner beroende på celltyp. Följande är några av dem:
Turgor
Cellturgor är nivån av hydrostatiskt tryck mot cellväggen i växtcellen. Detta tryck är under kontroll av vakuoler, som får olika ämnen inuti, inklusive joner, för att producera variabla inre osmotiska miljöer jämfört med cytosolens. Den olika osmolariteten på båda sidor av vakuolmembranet gör att vattnet passerar membranet, antingen inåt eller utåt. De ämnen som bidrar till vakuol-osmolariteten kan korsa vakuolmembranet genom ATP-beroende transport medierad av jonpumpar. H(+)-ATPas och H ( + ) – pyrofosfatas kan bilda protongradienter mellan båda sidor av vakuolmembranet, och dessa gradienter används för att transportera andra molekyler. Förmågan att lagra vatten inuti vakuolen är avgörande för växtcell växa efter mitos. Växtceller kan öka sin storlek 10 till 20 gånger, vilket är mycket användbart för kroppsväxten att växa och för att modifiera formen på växtorganen. Tillväxten medierad av hydrostatiskt tryck sparar energi eftersom det är billigare att öka mängden vatten än att syntetisera nya molekyler (djurcellstillväxt baseras på molekylär syntes). Det är säkrare för växtceller att ackumulera vatten i vakuolen eftersom de cytosoliska molekylerna på detta sätt inte späds ut, vilket skulle äventyra cellöverlevnaden.
Lagring
vakuoler är den sista stationen för vissa vesikulära trafikvägar. I vissa celler är de facket för att lagra kolhydrater och proteiner. Detta händer tydligt i frön, där vakuoler ackumulerar proteiner som behövs under spiring. Lagringsvakuoler blir lytiska vakuoler under celldifferentiering. Till skillnad från djur har växter inte ett utsöndringssystem, och de kan inte heller röra sig för att undvika giftiga ämnen. I växter lagras potentiellt farliga ämnen i vakuoler. På detta sätt finns metabolismrester och giftiga ämnen som tungmetaller (kadmium, zink och nickel) i vakuoler. Dessutom lagrar de också andra ämnen som pigment (till exempel antocyaniner) i epidermala celler av kronblad, giftiga ämnen mot växtätare, hartser, alkaloider som opium etc. Det mesta av smaken av frukt och grönsaker är resultatet av ämnen som lagras i vakuoler.
Nedbrytningscentra
lytiska vakuoler finns i vegetativa vävnader, så de är också kända som vegetativa vakuoler. De innehåller enzymer som proteaser och nukleaser, liksom ett antal proteiner som är involverade i försvaret mot patogener. Protonpumpar som sätts in i vakuolmembranet går in i protoner i vakuolen och surgör det inre innehållet. Det låga pH-värdet och de lytiska enzymerna tillåter nedbrytningsprocesser. Vakuoler har en liknande roll som lysosomer av djurceller. Dessutom, som lysosomer, deltar vakuoler i autofagi. Vakuolära bearbetningsenzymer är proteiner som också finns i vakuoler. De omvandlar molekylära prekursorer som anländer till vakuolen som inaktiva molekyler till aktiva molekyler.
apoptos
vakuoler är involverade i växtcell apoptos via en mekanism som kallas autolys. Dessutom förekommer en typ av celldöd som kallas överkänslig celldöd i växtceller när vakuolmembranet bryts.
andra
det finns specialiserade vakuoler i olika växtvävnader. Till exempel, i fröets inre tegument, ackumulerar vakuoler flavonoider för skydd mot ultraviolett ljus. Flavonoider syntetiseras i den cytosoliska ytan av endoplasmatiska retikulummembran och translokeras sedan till det inre av vakuoler för en slutlig kemisk bearbetning. I vakuolmembranet finns transportörer för att utföra denna translokation.
vissa växtarter, som brassicas, har vakuoler i sina vegetativa vävnader för att upphäva växtätare. Dessa vakuoler lagrar proteiner, såsom myrosinaser. När de väl släppts av växtätaraktiviteten bryter dessa enzymer ned molekylära föreningar av bladen som blir giftiga för djuret. Celler som lagrar myrosin är kända som myrosinceller och kan hittas nära bladets vaskulära buntar.
växter saknar immunförsvar så att varje cell har sitt eget försvarssystem. Försvarsproteiner och enzymer finns i vakuoler. Det finns två försvarsmekanismer som vakuoler kan utföra (Figur 3): vakuolmembran kollapsar och fusion av membranvakuol med plasmamembran. Virala infektioner leder till vakuolmembranbrott och frigör enzymer i cytosolen, där de kan attackera virus. Fusionen av vakuolmembran och plasmamembran frigör vakuolenzymer till det extracellulära utrymmet där de kan döda patogener som bakterier.
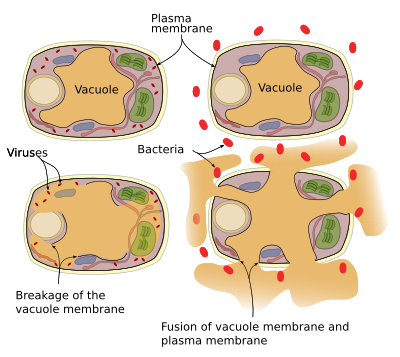
vesikulär handel
vakuoler är en del av vesikulär trafik. I själva verket kan de betraktas som en slutprodukt av vesikulär handel eftersom deras bildning och underhåll beror på de inkommande vesiklarna. Molekyler som kommer att lagras eller brytas ned, inkluderade hydrolytiska enzymer, liksom alla membranmolekyler är riktade mot vakuoler via vesiklar. Molekyler kan följa olika vesikulära vägar för att komma till vakuoler:
endoplasmatisk retikulum> Golgi-apparat> vakuol; Golgi-apparat> Pre-vakuolär fack > vakuol. Detta är standardvägen för att transportera hydrolytiska enzymer mot vakuoler. Pre-vakuolära fack liknar multivesikulära kroppar / sena endosomer av djurceller. Kärleksfullt väljs inte hydrolytiska enzymer i Golgi-apparaten av 6-fosfat-mannosdelar, utan av en sekvens av aminosyror belägna i deras aminosyrakedja. Det finns specifika sekvenser av aminosyror för att rikta proteiner till de hydrolytiska vakuolerna och andra sekvenser är specifika för att flytta andra proteiner mot lagringsvakuolerna. Alla proteiner riktade mot vakuoler har en signalsekvens, och de måste erkännas specifikt av receptorer.
endoplasmatisk retikulum> vakuol. Molekyler kan komma till vakuoler direkt från endoplasmatisk retikulum. Denna väg är framträdande i frön som en väg för lagring. Men i andra växtceller, som i löv, kan denna väg vara mycket sällsynt. Vesiklar som reser från endoplasmatisk retikulum till vakuoler är oberoende av COP-II-rockar, som behövs för vesiklar riktade mot Golgi-apparaten. I endoplasmatisk retikulum-vakuolväg finns det ibland mellanliggande fack, men de är övergående membranbundna organeller där molekyler hålls kort innan de anländer till vakuolen. Denna vesikulära väg kan härledas från autofagicellulära komponenter.
plasmamembran > vakuol. Endocytiska vesiklar smälter direkt med vakuoler, som fungerar som tidiga endosomer.
Bibliografi
Marty F. 1999. Plantera vakuoler. Växtcell 11:587-600.
Pereira C., Pereira S, Pissarra J. 2014. Leverans av proteiner till växtvakuolen – en uppdatering. Internationell tidskrift för molekylära vetenskaper 15: 7611-762.
Shimada T, Takagi J, Ichino T, Shirakawa M, Hara-Nishimura I. 2018. Växt vacuole. Årlig granskning i växtbiologi. https://doi.org/10.1146/annurev-arplant-042817-040508.
Taiz L. 1992. Anläggningen vacuole. Tidskrift för Experimentell Biologi 172: 113-122.
Zhang C, Hicks G R, Raikhel NV. 2014. Växtvakuolmorfologi och vakuolär handel. Gränser i Växtvetenskap 5: 476. ![]()